Расчет равновесного потенциала для ионов 11 страница
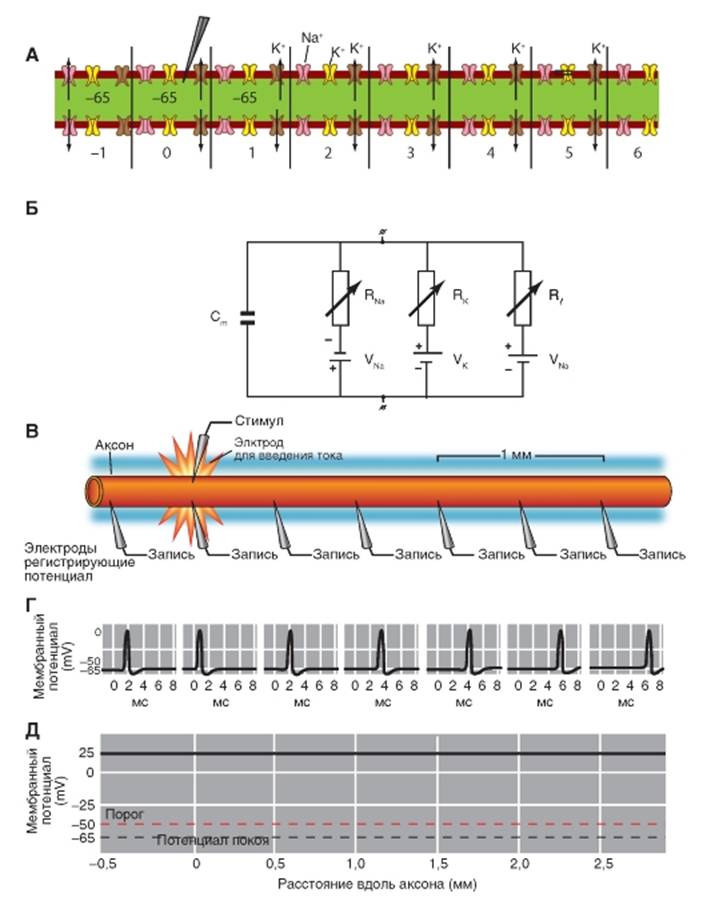
На рисунке 1-86 А представлена схема немиелинизированного волокна, для удобства обсуждения механизма распространения потенциала действия разбитая на фрагменты, каждый из которых включает один потенциалуправляемый Na+-канал (показан розовым цветом), находящийся в состоянии покоя, один потенциалуправляемый К+-канал (показан желтым цветом), находящийся в состоянии инактивации, и один канал утечки (показан коричневым цветом), через который осуществляется выход ионов К+ во внешнюю среду.
Для понимания механизмов распространения потенциала действия необходимо учитывать как пассивные (емкость и сопротивление), так и активные (активация потенциалупраляемых каналов) свойства мембраны нервного волокна. Рассмотрим эквивалентную электрическую схему (рис. 1-86 Б), представляющую собой электрическую модель нервного волокна, удобную для рассматривания проведения потенциала действия. Сопротивлением аксоплазмы и наружной среды можно в определенном смысле пренебречь. На этой схеме представлен лишь один элемент мембраны.
Каждый элемент имеет емкость мембраны Сm (рис. 1-86 Б), сопротивление потенциалуправляемого Na+-канала (RNa), натриевую батарею VNa, сопротивление потенциалуправляемого К+-канала (RK, калиевую батарею сопротивление канала утечки, К2Р-канала (Rl), батарею канала утечки Vl. Сопротивление канала утечки и его батарея приведены для демонстрации движения ионов по круговым локальным токам через эти каналы в невозбужденных участках мембраны аксона, соседствующих с возбужденным.
|
|
|
Пространственное распределение потенциала по мембране определяется токами, протекающими через нее. Однако при распространении потенциала действия по аксону ток зависит от координаты (эта пространственная неоднородность зависит, например, от наличия открытых потенциалуправляемых каналов). Именно поэтому локальное изменение потенциала (его первая производная) определяется локальными токами (т.е. ток как функция координаты). В свою очередь, локальный ток через мембрану определяется локальным потенциалом, и изменение тока (его первая производная) зависит как от пространственного изменения потенциала (резистивный компонент), так и от временного изменения потенциала (емкостной компонент).
На рисунке 1-86 В показан аксон с введенным в него стимулирующим электродом и группой регистрирующих микроэлектродов. Потенциалы действия в каждой регистрируемой точке показаны на рис. 1-86 Г и измеряются регистрирующими электродами у основания аксона (V1), а также в различных точках вдоль него (V2, V3, V4, V5, V6); график этих изменений представлен на рис. 1-86 Д. Уже из этих последних рисунков видно, что при распространении потенциала действия по аксону ни его амплитуда, ни его форма не меняются. Механизм явления представлен на следующем рисунке.
|
|
|
Рис. 1-86. Распространение потенциала действия на примере немиелинизированного волокна.
А - аксон в состоянии покоя. Потенциалуправляемые Na+-каналы (показаны розовым цветом) находятся в состоянии покоя, потенциалуправляемые К+-каналы (показаны желтым цветом) находятся в состоянии инактивации, работают только каналы утечки (показаны коричневым цветом), через которые осуществляется выход ионов К+ во внешнюю среду. Б - электрические свойства аксона можно моделировать, используя радиоэлектронные элементы. Показана эквивалентная электрическая схема одного фрагмента мембраны аксона, где Cm - емкость мембраны, RNa - сопротивление потенциалуправляемого Na+-канала, VNa - натриевая батарея, RK - сопротивление потенциалуправляемого К+-канала, VK - калиевая батарея, Rl - сопротивление канала утечки (К2Р-канала), Vl - батарея канала утечки. Мембрана немиелинизированного нервного волокна напрямую контактирует с внешней средой и, таким образом, в покое обмен ионами между внутри- и внеклеточной средами (ионные токи через плазматическую мембрану, прежде всего по каналам утечки) может происходить в любой точке немиелинизированного нервного волокна. В - через стимулирующий электрод подается пороговый импульс электрического тока, вызывающий возникновение потенциала действия. Г - потенциал действия распространяется по волокну без затухания и измеряется регистрирующими электродами у основания аксона (V1), а также в различных точках вдоль него (V2, V3, V4, V5, V6). Д - график, демонстрирующий неизменную амплитуду потенциала действия на любом удалении от точки стимуляции
|
|
|
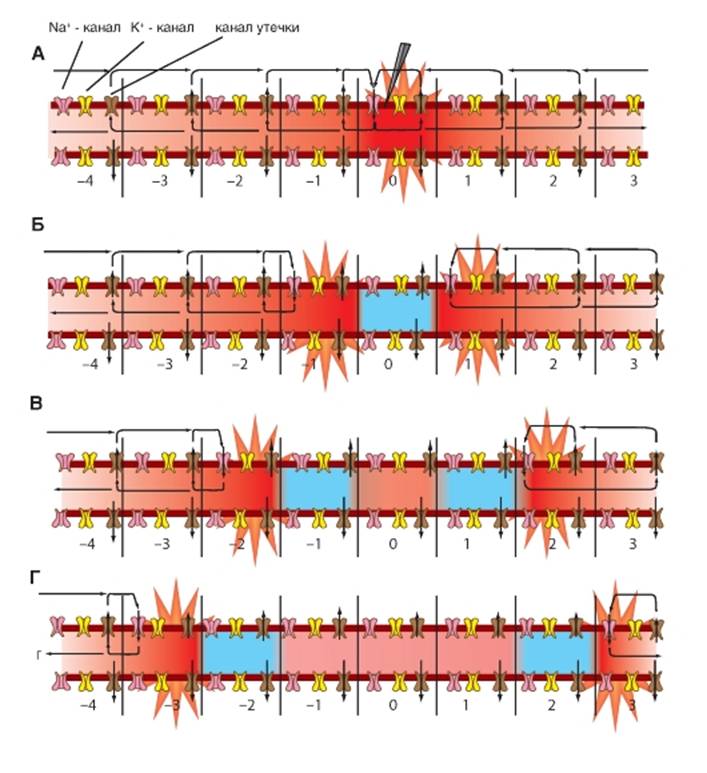
Механизм распространения потенциала действия по немиелинизированному волокну
Рис. 1-87. Механизм проведения потенциала действия в немиелинизированном волокне.
А - схема немиелинизированного нервного волокна в покое. Для описания механизма проведения возбуждения по немиелинизированному нервному волокну аксон разбит на условные фрагменты 0, 1, 2, 3...n, каждый из которых содержит потенциалуправляемые Na+- и К+-каналы и каналы утечки. Зеленый цвет всех фрагментов аксона соответствует состоянию покоя (рис. 1-86 А). А - раздражение фрагмента (0) нервного волокна приводит к открытию потенциалуправляемых Na+-ионных каналов, что ведет к возникновению входящего Na+-тока (фрагмент волокна окрашен красным цветом) и генерации фазы деполяризации потенциала действия на этом участке аксона. Поскольку внутриклеточная среда служит проводником, входящий в зону (0) ток распространяется в обе стороны от возбужденного участка. Так как мембрана нервного волокна не идеальный изолятор, распространяющиеся токи будут покидать волокно через единственно открытые каналы утечки, т.е. возникнут выходящие токи, протекающие через соседние участки мембраны (зоны 1, а также зоны 2, 3 и т.д.). Таким образом, локальный входящий ток всегда вызывает выходящие токи в прилегающих невозбужденных участках, т.е. возникают локальные круговые токи между возбужденным и прилегающими невозбужденными участками. Линии показывают локальные круги тока, текущего между деполяризованной и недеполяризованной областями внутри и снаружи аксона и через зоны мембраны, находящиеся в покое (зоны 1), а также на большем удалении. Б - токи, протекающие через зоны (1), электротонически изменяют их потенциал, приводя к разряду мембранной емкости и, следовательно, к деполяризации. Важно отметить, что эта фаза распространения потенциала действия опосредована пассивными свойствами мембраны нервного волокна (аналогично распространению пассивного электротонического потенциала). Однако в отличие от пассивного электротонического потенциала, деполяризация, возникающая в зонах (1), при распространении потенциала действия достигает порога открывания потенциалуправляемых Na+-каналов. Это приводит к лавинообразному открытию всех Na+-каналов в зонах (1), что сопровождается возникновением фазы деполяризации потенциала действия в этих зонах волокна. Входящий Na+-ток вызывает локальные круговые токи между возбужденными зонами (1) и прилегающими участками - невозбужденными зонами (2), а также зоной 0. Несмотря на то что в результате этого процесса зона (0) также частично деполяризуется, потенциалуправляемые Na+-каналы в ней находятся в состоянии инактивации, а вся зона - в состоянии рефрактерности (голубой цвет на рисунке). Именно поэтому здесь потенциал действия вновь не возникнет. В зонах (2), находящихся в состоянии покоя, локальные круговые токи электротонически сдвигают потенциал покоя в сторону деполяризации. Когда деполяризация достигает уровня критического потенциала, активируются потенциалуправляемые Na+-каналы и соответственно появляется входящий Na+-ток, формирующий в зонах (2) фазы деполяризации потенциалов действия. В - далее входящий Na+-ток, протекающий через мембрану в зонах 2, вызывает локальные круговые токи между возбужденными зонами 2 и невозбужденными зонами 3, а также зонами 1 и 0. Стрелки вновь показывают локальные круги тока. Хотя в результате этого процесса зоны 1 также частично деполяризуются, потенциалуправляемые Na+-каналы в них находятся в состоянии инактивации. Там регистрируются фазы реполяризации потенциала действия, а сами зоны находятся в состоянии рефрактерности (голубой цвет на рисунке). Именно поэтому в зонах 1 потенциал действия не возникнет. Его также не будет в зоне 0, поскольку она удалена от центров возбуждения. Вспомним, что пассивный электротонический потенциал распространяется с затуханием. Таким образом, электротоническая деполяризация, вызванная круговыми токами между возбужденными зонами 2, а также зонами 1 и 0, не достигает порога активации потенциалуправляемых Na+-аналов в зонах 1 и 0, и там потенциалы действия не возникают. В зонах 3 потенциал действия будет вызван. Г - Входящий Na+-ток, протекающий через мембрану в зонах 3, вызывает локальные круговые токи между возбужденными зонами 3 и невозбужденными зонами 4, а также зонами 2, 1 и 0. Стрелки вновь показывают локальные круги тока. Хотя в результате этого процесса зоны 2 также частично деполяризуются, потенциалуправляемые Na+-каналы в них находятся в состоянии инактивации. Там регистрируются фазы реполяризации потенциала действия, а сами зоны находятся в состоянии рефрактерности (голубой цвет на рисунке). Именно поэтому в зонах 2 потенциал действия не возникнет. Не возникнет потенциал действия и в зонах 1 и 0, поскольку они удалены от центров возбуждения. Вспомним опять, что пассивный электротонический потенциал распространяется с затуханием. Таким образом, электротоническая деполяризация, вызванная круговыми токами между зонами 3 и 2, 1, 0, не достигает порога активации потенциалуправляемых Na+-каналов, и потенциал действия там не возникает. В зонах 4 потенциал действия будет вызван
|
|
|
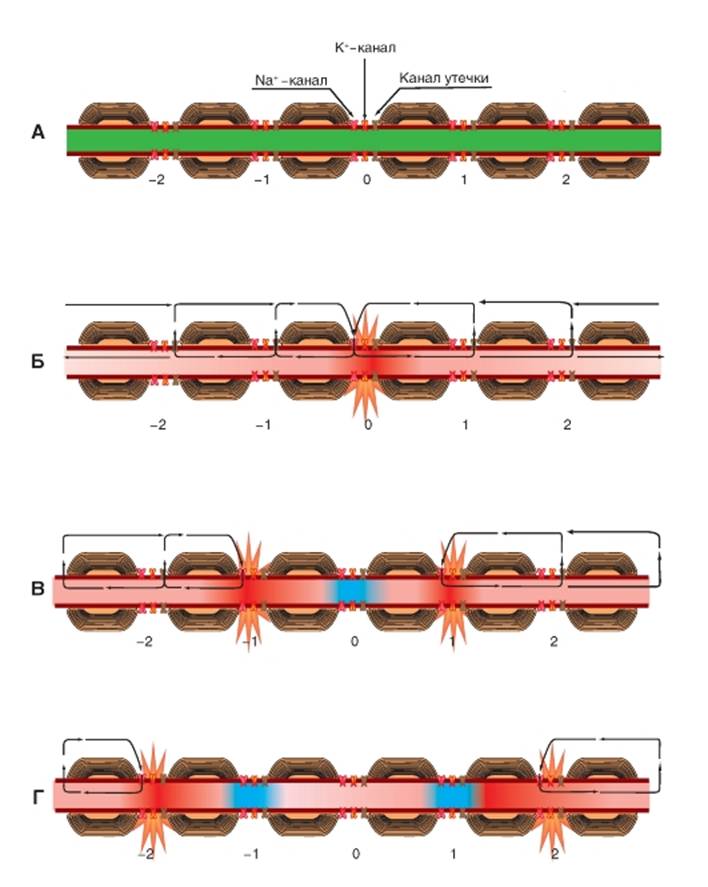
Механизм распространения потенциала действия по миелинизированному волокну
Рис. 1-88. Механизм проведения потенциала действия в миелинизированном волокне.
А - для описания механизма проведения возбуждения по миелинизированному нервному волокну аксон разбит на условные фрагменты 1, 2, 3...n, каждый из которых содержит 1 перехват Ранвье с потенциалуправляемыми Na+- и К+-каналами и каналами утечки. Внешняя поверхность мембраны перехватов Ранвье заряжена положительно, а внутренняя поверхность их мембраны заряжена отрицательно. Остальные области мембраны покрыты миелином. Зеленый цвет всех фрагментов аксона соответствует состоянию покоя. Электрическая активность в миелинизированном аксоне возможна только в области перехватов Ранвье, где ионы могут проходить через мембрану. Эти регионы мембраны имеют большую плотность потенциалуправляемых Na+-каналов. Участки нервного волокна, которые покрыты миелином, имеют значительно более высокое сопротивление, чем обычная плазматическая мембрана. Таким образом, локальные круговые токи, которые необходимы для генерации потенциала действия, на миелинизированных участках не возникают. Потенциалы действия возникают только в перехватах Ранвье. Миелинизированное волокно представляет собой значительно более качественный кабель по сравнению с немиелинизированным аксоном. Как следствие, электрические сигналы будут распространяться электротонически на большие расстояния. Б - схема состояния миелинизированного нервного волокна при действии на перехват Ранвье (0) порогового раздражителя. Вспомним, что у миелинизированных нервных волокон, вследствие высокой концентрации свободных ионов внутри и снаружи нервного волокна, внутри- и внеклеточная среды являются хорошими проводниками. Кроме того, ток может протекать через мембрану только в районах перехватов Ранвье. Раздражение перехвата Ранвье (0) нервного волокна приводит к открытию потенциалуправляемых Na+-ионных каналов, что ведет к возникновению входящего Na+-тока и генерации фазы деполяризации потенциала действия. В результате этого между возбужденным (0) и соседними перехватами Ранвье (1) и (2) возникает разность потенциалов, что вызывает локальный круговой ток, текущий между деполяризованной и недеполяризованной областями внутри аксона и замыкающегося через экстрацеллюлярную жидкость. Стрелки показывают локальный круг тока, текущего между деполяризованной и недеполяризованной областями внутри и снаружи аксона, проникая через покоящуюся зону перехвата Ранвье (1) мембраны и частично на большем удалении. В - раздражение перехватов Ранвье (1) нервного волокна приводит к открытию потенциалуправляемых Na+-ионных каналов, что ведет к возникновению входящего Na+-тока и генерации фазы деполяризации потенциалов действия на этих участках. В результате между возбужденными перехватами Ранвье (1) и между соседними перехватами Ранвье (2) возникает разность потенциалов, что вызывает локальные токи, текущие между деполяризованной и недеполяризованной областями. Такой же локальный ток течет и между перехватами Ранвье (1) и перехватом Ранвье (0), но, поскольку последний находится в стадии рефрактерности (Na-каналы находятся в состоянии инактивации и не могут быть открыты), перехват Ранвье (0) возбудиться не может. Г - раздражение перехватов Ранвье (2) нервного волокна приводит к открытию потенциалуправляемых Na+-ионных каналов, что ведет к возникновению входящего Na+-тока и генерации фазы деполяризации потенциалов действия (нижняя часть рисунка) на этих участках. В результате между возбужденными перехватами Ранвье (2) и соседними перехватами Ранвье (3) возникает разность потенциалов, что вызывает локальные токи, текущие между деполяризованной и недеполяризованной областями. Такой же локальный ток течет и между перехватами Ранвье (1) и перехватом Ранвье (0), но, поскольку перехваты Ранвье (1) находятся в стадии рефрактерности (Na-каналы находятся в состоянии инактивации и не могут быть открыты), они возбудиться не могут
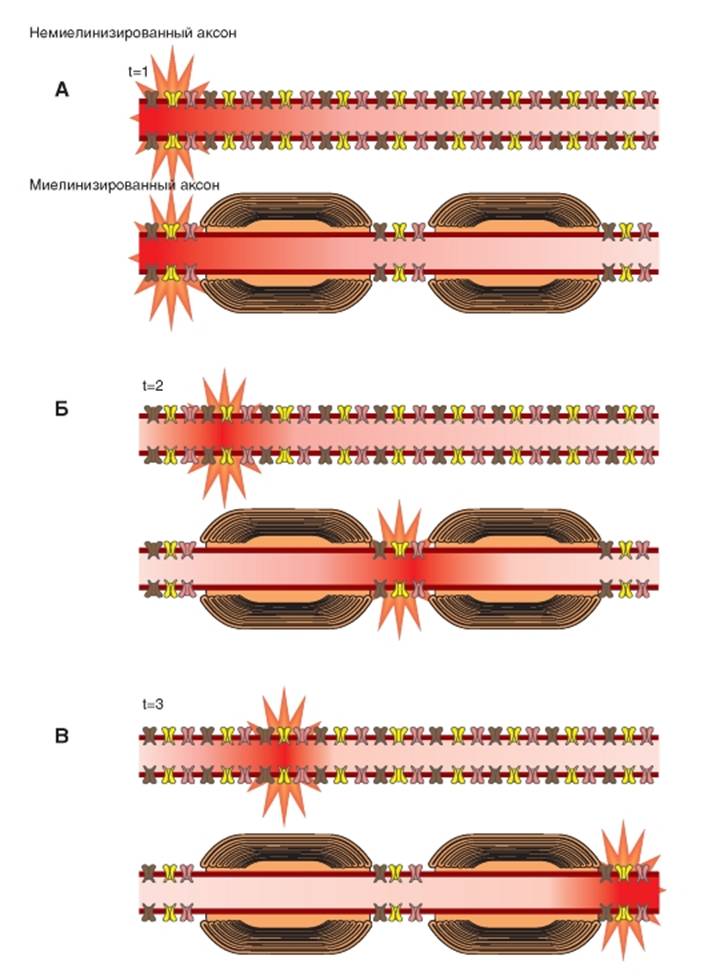
Проведение возбуждения по двум типам волокон
Распространение возбуждения в немиелинизированном и миелинизированном нервном волокне суммарно и схематически представлено на рис. 1-89. В отличие от немиелинизированного волокна, возбуждение передается от одного перехвата Ранвье к другому, и потенциалы действия возникают только в этих зонах. То есть происходит «перескакивание» возбуждения от одного перехвата Ранвье к другому. Такой механизм, характерный только для миелинизированных нервных волокон, называется сальтаторным проведением. Электрический ток течет через окружающую экстрацеллюлярную жидкость снаружи миелинового покрытия, так же как и через аксоплазму аксона от перехвата к перехвату, успешно возбуждая перехваты один за другим. Таким образом, нервный импульс «прыгает» по волокну, обусловливая наличие такого термина как «сальтаторное проведение». Пусть возбуждение возникло одновременно на двух типах волокон (рис. 1-89 А). Благодаря сальтаторному проведению у миелиновых волокон потенциал действия возникает сразу же на расстоянии в следующем перехвате Ранвье, тогда как в немиелинизированном волокне потенциал действия возникает только на соседнем участке (рис. 1-89 Б). Далее у миелиновых волокон потенциал действия возникает на следующем перехвате Ранвье, тогда как в немиелинизированном
волокне потенциал действия возникает только на следующем участке (рис. 1-89 В).
Сальтаторное проведение ценно по двум причинам. Во-первых, вызывая деполяризацию только перехватов Ранвье, возбуждение «прыгает» вдоль длинных участков оси нервного волокна, и этот механизм увеличивает скорость проведения возбуждения примерно в 5-50 раз. Во-вторых, сальтаторное проведение консервирует энергию у аксона, потому что деполяризуются только перехваты, позволяя при генерации потенциалов действия терять в 100 раз меньше ионов и, таким образом, требуя минимального метаболизма для восстановления натриевой и калиевой разностей концентраций относительно мембраны после серии нервных импульсов.
Другая любопытная черта сальтаторного проведения в больших миелиновых волокнах заключается в следующем. В конце потенциала действия, когда потенциалуправляемые Na+-каналы инактивируются, реполяризация развивается так быстро, что многие из К+-каналов даже не успевают открыться. Таким образом, проведение нервного импульса в миелинизированных волокнах опосредовано в основном ионной проводимостью через потенциалуправляемые Na+-каналы и каналы утечки. Вклад потенциалуправляемых К+-каналов незначителен. Этим объясняется тот факт, что потенциалы действия миелинизированных нервных волокон часто не имеют фазы следовой гиперполяризации.
Рис. 1-89. Сравнение общих принципов проведения возбуждения по немиелинизированному и миелинизированному нервному волокну
Внеклеточные потенциалы
В качестве предварительного усилителя обычно применяют усилитель постоянного тока с дифференциальным входом (рис. 1-90 А). Это связано с тем, что измеряется разность потенциалов между возбужденными и невозбужденными участками нерва.
Далее (рис. 1-90 Б) показаны записи внеклеточного потенциала нервного волокна на примере ответов седалищного нерва лягушки. Потенциал действия нерва во многом отличается от потенциала действия отдельных волокон. Он не подчиняется закону «все или ничего», поскольку при увеличении силы раздражающего импульса увеличивается число возбужденных аксонов и, следовательно, растет суммарная амплитуда самого потенциала действия. Сложный потенциал действия нерва отличается также своими порогом, формой и рядом других характеристик.
По скорости проведения возбуждения, длительности различных фаз потенциала действия и строению нервные волокна принято выделять три основных типа, обозначаемых буквами А, В и С (рис. 1-90 В).
Волокна типа А делятся на четыре подгруппы. Это α, β, γ и δ. Все они покрыты миелиновой оболочкой. Наиболее толстые из них, так называемые альфа-волокна (аА), у теплокровных животных и человека имеют диаметр 12-22 μм и характеризуются значительной скоростью проведения возбуждения: 70-120 м/с. Такие волокна проводят возбуждение от моторных нервных центров спинного мозга к скелетным мышцам и от рецепторов мышц к соответствующим нервным центрам. Пик потенциала действия аА волокон у теплокровных длится 0,4-0,5 мс. После его окончания развивается следовая деполяризация, которая продолжается 15-20 мс и переходит в следовую гиперполяризацию длительностью около 40-60 мс.
Три другие группы волокон типа А (β, γ и δ) имеют меньший диаметр, меньшую скорость проведения и более длительный потенциал действия. Это преимущественно чувствительные волокна, проводящие возбуждение от различных рецепторов в центральную нервную систему. Исключение составляют γА-волокна, значительная часть которых проводит возбуждение от клеток спинного мозга к так называемым интрафузальным мышечным волокнам, входящим в состав рецепторов мышц - мышечных веретен (см. раздел «Мышцы»).
Дата добавления: 2018-08-06; просмотров: 323; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!