Расчет равновесного потенциала для ионов 9 страница
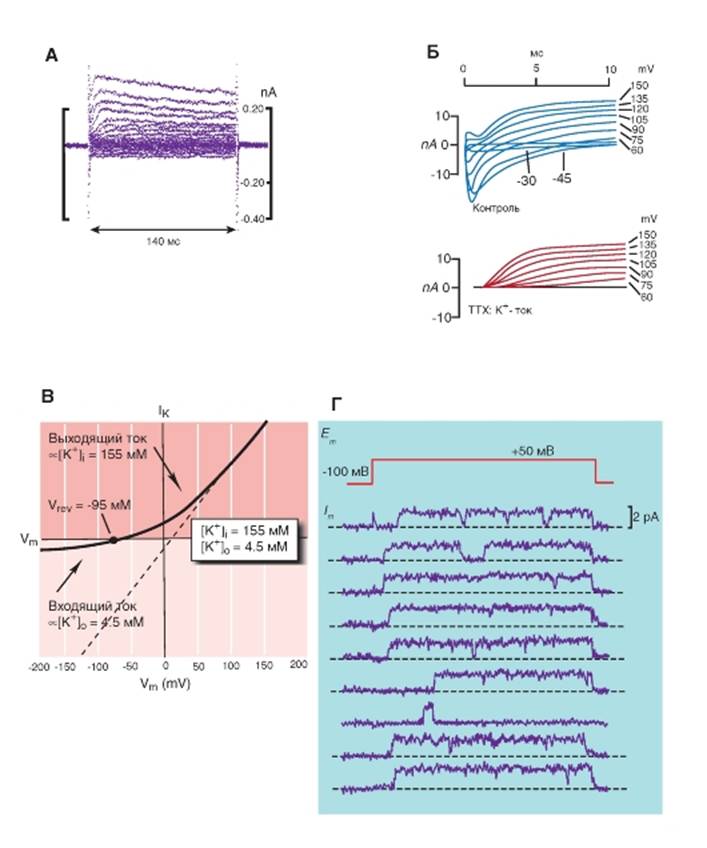
Обсудим некоторые данные, характеризующие работу абстрактных К+-каналов. На рисунке 1-74 В представлена вольтамперная кривая K+-каналов, рассчитанная при помощи уравнения Гольдмана- Ходжкина-Катца. Сплошная кривая отражает K+- ток, предсказанный на основании уравнения с допущением, что мембрана хорошо проницаема для ионов K+, причем [K+]in равна 155 мМ, а [K+]out равна 4,5 мМ. Величина -95 мВ - потенциал реверсии (reversal potential: Vrev), потому что при этом потенциале направление тока меняет значение, а в самой точке, равной -95 мВ, ток равен нулю. В физике направление тока всегда такое же, как и направление перемещения положительных зарядов. Ток ионов, текущий в клетку, будет, таким образом, называться inward current, а ток ионов, текущий из клетки, - всегда outward. Пунктирная линия демонстрирует ток, который можно ожидать, если [K+]in равна [K+]out и соответствует 155 мМ.
Наконец на рис. 1-74 Г показана регистрация одиночного канала К+-канала.
Традиционно K+-токи и соответственно K+-каналы вследствие их функции были описаны и классифицированы главным образом в возбудимых клетках. Они включали классические:
• K+-каналы задержанного выпрямления (delayed outward rectifier K+-channels (currents) - IK);
• быстрые транзиторные K+-каналы выходящего тока (fast transient K+-channels (currents) - IA, или transient outward current - Ito);
• K+-каналы аномального выпрямления с током входящего направления (inward rectifier K+- channels - IK1);
• Ca2+-активируемые K+-каналы (Ca2+ dependent K+-channels - KCa);
|
|
|
• каналы, через которые проходит ток утечки (leak currents), определяемый преимущественно ионами K+.
Чуть больше десятилетия назад была принята первая стандартная номенклатура для генов шести трансмембранных сегментов потенциалуправляемых K+каналов, названных KV-системой. В нее прежде всего вошли:
• K+-каналы задержанного выпрямления (delayed outward rectifier K+-channels - IK);
• быстрые транзиторные K+-каналы выходящего тока (fast transient K+-channels (currents) - IA, или transient outward current - It0).
Эта номенклатура была основана на выведенных филогенетических связях. Каналы, в составе которых обнаружено более 65% идентичных аминокислотных остатков, были объединены в одно подсемейство. В отдельную группу были выделены K+-каналы аномального выпрямления с током входящего направления (inward rectifier K+-channels), которая была названа Kir.
Таким образом, принятая в настоящее время классификация делит K+-каналы на:
• потенциалуправляемые К+-каналы (KV), которые включают:
- K+-каналы задержанного выпрямления (delayed outward rectifier K+-channels - IK);
- быстрые транзиторные ^-каналы выходящего тока (fast transient K+-channels (currents) - IA, или transient outward current - Ito);
• K+-каналы аномального выпрямления с током входящего направления (inward rectifier K+-channels - Kir);
• Ca2+-активируемые K+-каналы (KCa);
|
|
|
• K+-каналы с двумя петлями в домене (two-P K+-channels - К2Р).
Рис. 1-74. Примеры оригинальной регистрации whole-cell К+-тока, вольтамперной характеристики whole-cell К+-тока и активности одиночного канала
Характеристики К+-токов
Разлагая на компоненты ионные токи можно вычленить входящий Na+-ток и выходящий К+-ток, представив их отдельно. Анализ этих «макроскопических токов», т.е. токов, зарегистрированных методом voltage-clamp или patch-clamp в конфигурации whole-cell,обсуждался ранее. Этот анализ позволяет разделить К+-токи на потенциалуправляемые (включающие К+-токи задержанного выпрямления и быстрые транзиторные К+-токи выходящего направления), К+-токи аномального выпрямления входящего направления, Ca2+-активируемые К+-токи и токи утечки. Но детальную информацию дал анализ одиночных К+-каналов методом patch-clamp.Микроскопические механизмы, то есть механизмы на уровне токов, текущих через одиночные каналы, лежащие в основе макроскопических вольт-амперных характеристик, то есть процессов, происходящих на уровне токов, текущих через целую клетку.
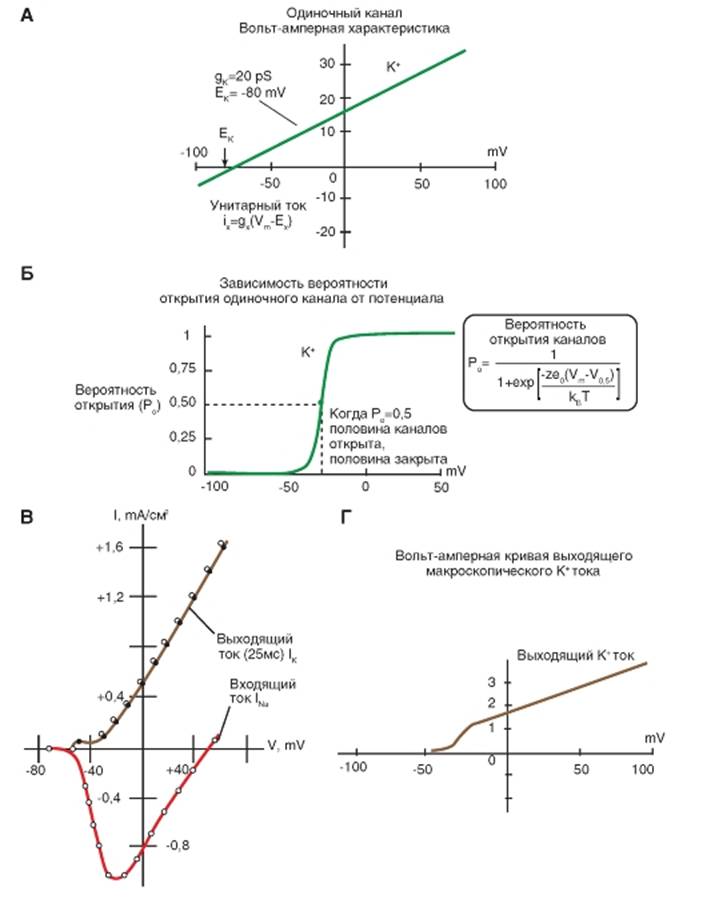
На рис. 1-75 А линия обозначает вольт-амперную зависимость идеализированного открытого калиевого канала. Т.к. мы исходим из начальной предпосылки, что канал в нашем случае будет полностью открыт все время (т.е. что проводимость каналов не будет функцией напряжения) тогда ток, текущий через них, будет линейным или «омическим».
|
|
|
На рис. 1-75 Б представлена кривая, которая показывает вероятность того, что калиевый канал будет находиться в открытом состоянии (вероятность открытия калиевого канала). Уравнение, приведенное во вставке в рисунок, описывает приведенную кривую, если мы вставим значения zK = 5,3 и V0.5 = - 30 mV.
На рис. 1-75 В представлен отдельно макроскопический калиевый ток.
На рис. 1-75 Г представлена оценка макроскопического калиевого тока посредством умножения одноканальных токов из секции А рисунка на вероятность их открытия Ро из секции Б и на количество каналов (N). В данном случае мы устанавливаем количества каналов в 100 калиевых.
Рис. 1-75. Общая характеристика потенциалуправляемых K+-каналов иK+-токов. Описание в тексте
К+-каналы задержанного выпрямления
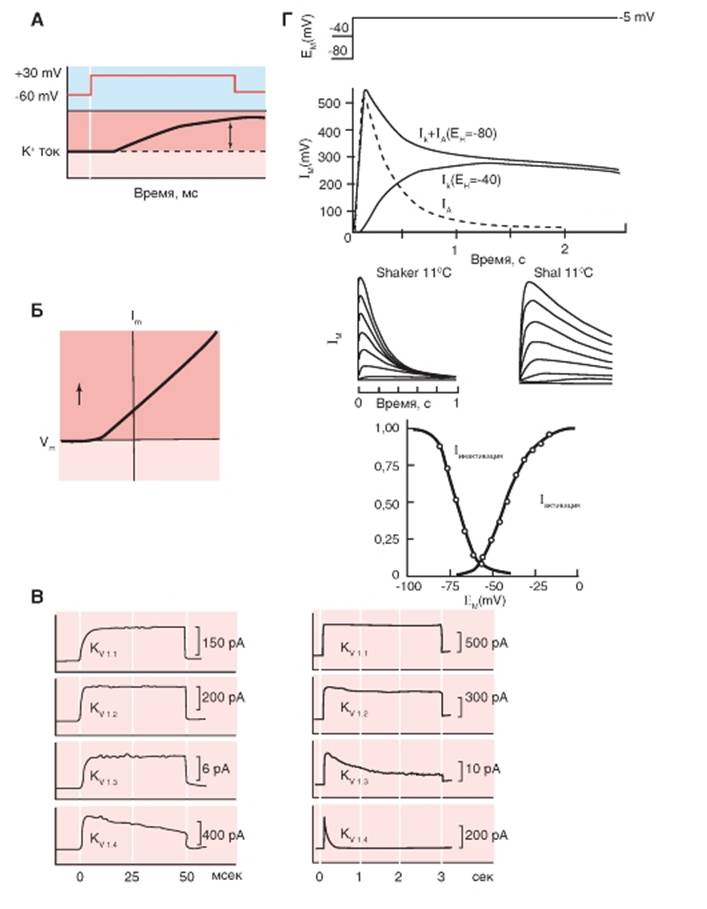
Обсудим некоторые данные, характеризующие работу первого типа потенциалуправляемых К+-каналов - К+-каналов задержанного выпрямления (delayed outward rectifier K+-channels - IK). Этот тип каналов отличается тем, что деполяризующие ступеньки активируют ток, но с задержкой, как это видно на рис. 1-76 А. Вольтамперная кривая, полученная на основе экспериментов, выполненных методомpatch-clamp в конфигурации whole-cell, однозначно демонстрирует наличие К+-каналов задержанного выпрямления (delayed outward rectifier K+-channels - IK). Ток течет только в выходящем направлении, как это показано на рис. 1-76 Б. Для оценки тока, текущего через каналы этого типа из клеток мозга млекопитающих, различные каналы (KV1.1,
|
|
|
KV1.2, KV1.3 и KV1.4) экспрессировали в ооциты ксенопуса. Результаты экспериментов, в которых потенциал смещался от -80 мВ до 0 мВ, показаны на рис. 1-76 В. Левая панель, представленная с большой степенью разрешения времени, демонстрирует, что некоторые из этих каналов активировались более медленно, чем другие. Правая панель, ужатая во времени, позволяет увидеть, что инактивация ускорялась от канала KV1.1 к каналу KV1.4.
Транзиторные К+-каналы
Данные, характеризующие работу второго типа К+-каналов, - быстрых транзиторных K+-каналов выходящего тока (fast transient K+-channels (currents) - IA, или transient outward current - It0) представлены на рис. 1-76 Г. Показаны результаты экспериментов, позволяющие построить кривую активации этого тока и его инактивации.
Рис. 1-76. Потенциалуправляемые К+-каналы (KV), относящиеся к типу К+-каналов задержанного выпрямления (delayed outward rectifier К"-channels - IK) и к типу быстрых транзиторных К+-каналов выходящего тока fast transient К+-channels currents - IA, илиtransient outward current - Ito
Кir-каналы
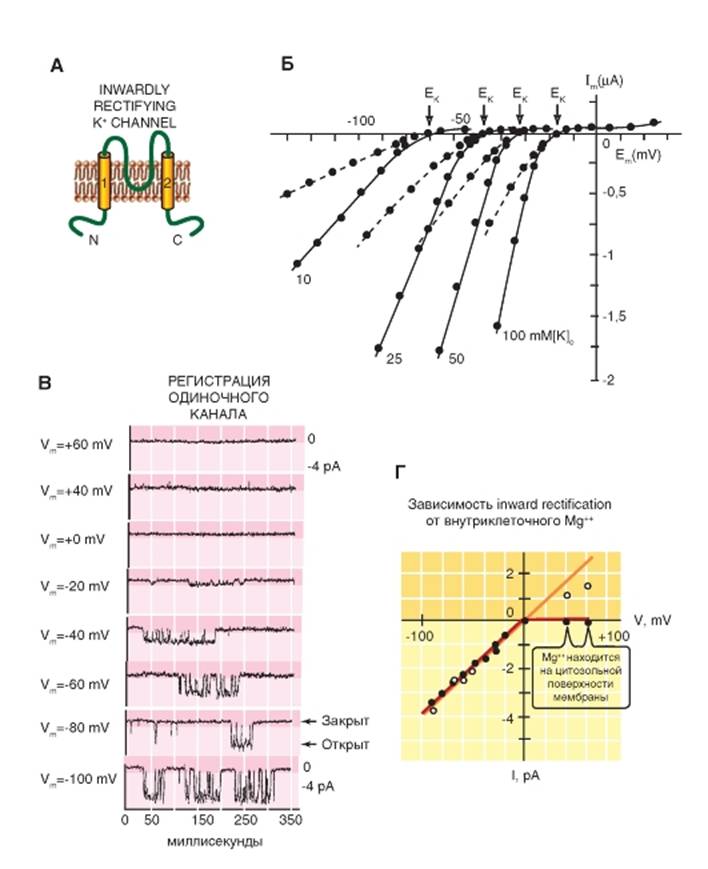
Класс трансмембранных белков с двумя сегментами включает 2S белки, образующие inward rectifiers channels (Kir), которые можно назвать «K+-каналами аномального выпрямления с током входящего направления». Терминали N- и C- этих каналов расположены в цитоплазме, P-петля между двумя трансмембранными сегментами формирует пору, и функционирующий
канал представляет собой тетрамер этих 2S/1P субъединиц (рис. 1-77 А). Вольтамперные характеристики при различных величинах [K+]0, характерные для этого типа каналов, представлены на рис. 1-77 Б. Там же показано значение ЕК для каждой величины [K+]o. На рисунке 1-77 В изображена зависимость открытия каналов от потенциала, а на рис. 1-77 Г представлена вольтамперная характеристика этого типа каналов.
Рис. 1-77. К+-каналы аномального выпрямления с током входящего направления (inward rectifier K+-chnnels - Kir)
Са2+-активируемые К+-каналы
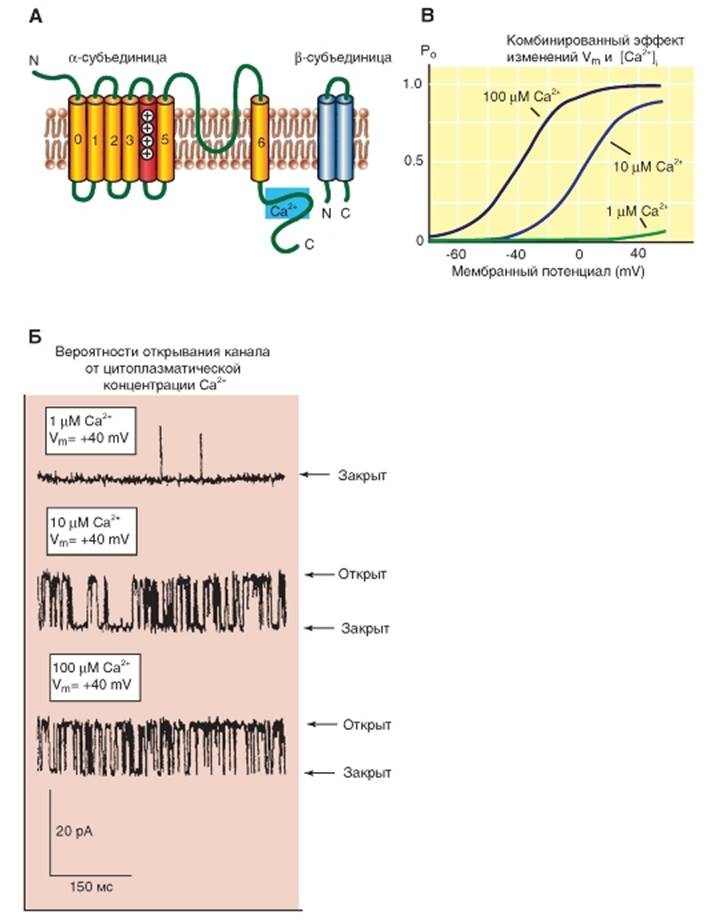
Еще один тип К+-каналов - так называемые Са2+-активируемые К+-каналы. Примером такого канала может служить, например, канал (Slo
или Slo1). Slo имеет 7S и кодирует высокопроводящий канал, Slo (0S-6S). В этом канале Р-область находится между SVI и SVII; функционирующий канал представляет собой тетрамер, но, в отличие от других каналов, Slo имеет N-терминаль, расположенную во внеклеточной среде (рис. 1-78 А).
Са2+-активируемые К+-каналы описаны достаточно давно и включают два подтипа. Одна из этих групп включает три KCa-канала с низкой проводимостью (KCa2.1, KCa2.2, и KCa2.3) и канал KCa3.1 с промежуточной проводимостью. Эти каналы непотенциалуправляемые и активируются низкими концентрациями внутриклеточного Ca2+ (<1,0 μΜ), в отличие от KCa1.1 (KCNMA1, Slo1), который активируется и потенциалом, и внутриклеточным Ca2+. Эти три KCa-канала с низкой проводимостью чувствительны к блокирующему действию апамина (100 пМ-10 нМ), что отличает их от всех других KCa-каналов. И KCa-каналы с низкой проводимостью, и каналы с промежуточной проводимостью играют важную роль во многих процессах, включающих Са2+-зависимый сигналлинг в электровозбудимых и электроневозбудимых клетках. Они не связывают свободные ионы Ca2+ непосредственно, а скорее, обнаруживают Ca2+ через кальмодулин, который постоянно связан с C-терминальным регионом. Связывание кальция с таким кальмодулином приводит к конформационным изменениям, ответственным за воротный механизм канала.
Вторая группа KCa-каналов включает KCа1.1 (Slo или Slo1), KCa4.1 (Slack или Slo2.2), KCa4.2 (Slick или Slo2.1) и KCa5.1 (Slo3). Каналы KCa1.1 изучали
в клетках мозга, улитки и мышцах, и известно, что варианты альтернативного сплайсинга их мРНК ответственны за их значительные функциональные различия. В отличие от KCa2 и каналов, связывание ионов Са2+ каналом KCа1.1 не зависит от его ассоциации с кальмодулином, но полагают, что определяется, по крайней мере, тремя связывающими двухвалентные катионы участками в цитоплазматической карбоксильной области каждой субъединицы канала. Два независимых высоко чувствительных Ca2+-связывающих участка, названных «calcium bowl», сформированы отрицательно заряженными сегментами в дистальной карбоксильной терминали белка и внутри первой RCK области (RCK: Regulator of Conductance for K+), кодирующей проксимальную C-терминаль белка. Третий низкоафинный связывающий двухвалентные катионы участок также найден в первой RCK области.
Хотя три других члена этой группы, KCa4.1, 4.2, и 5.2, были включены в KCa-спецификацию, так как все они являются членами этой структурно связанной группы генов, но в отличие от KCa1.1 активирующихся внутриклеточным Ca2+, ни один из вышеупомянутых каналов, повидимому, не может быть им активирован. Эти три нечувствительные к внутриклеточному Ca2+ канала активируются иначе. KCa4.2 и KCa4.1 активируются внутриклеточными ионами Na+ и Cl-, а KCa5.1 - внутриклеточным защелачиванием (сдвигом рН в щелочную сторону).
На рисунке 1-78 Б показаны токи через одиночные Са2+-активируемые К+-каналы, Вероятность открытия каналов значительно повышается с увеличением концентрации Са2+. На рис. 1-78 В показана зависимость вероятности открытия канала от потенциала на фоне различной концентрации Са2+.
Рис. 1-78. Са2+-активируемые K+-каналы (KCa)
Канал утечки - K2P
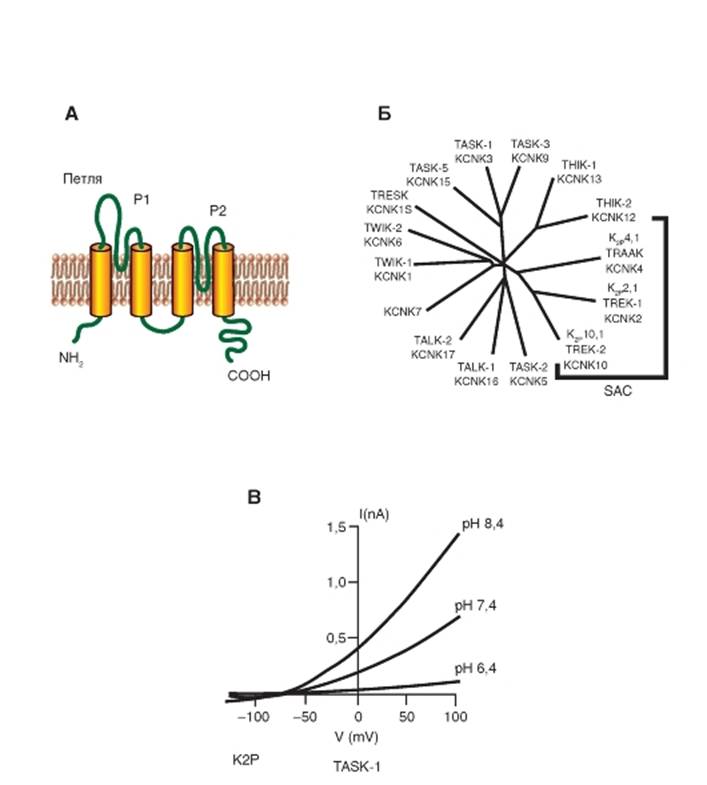
Каналы утечки представляют собой особую группу ионных каналов. Это одни из немногих каналов, о которых около 50 лет, т.е. со времени возникновения представлений о них, было мало что известно. Они имеют потенциалнезависимую основу и калиево-натриевую проводимость. В работах Б. Хилле (B. Hille) отмечено, что уже А. Ходжкин и А. Хаксли на фоне потенциала покоя демонстрировали небольшой компонент тока, приписываемый ими так называемому току утечки Il (leak current). Этот ток был небольшим и имел потенциалнезависимый механизм ионной проводимости. А. Ходжкин и А. Хаксли не установили ионную основу тока утечки. Было только показано, что на фоне потенциала покоя мембрана клетки относительно проницаема для ионов калия, выходящих из клетки, и крайне плохо (в 100 раз меньше) проницаема для ионов натрия, входящих в клетку. Эту проницаемость традиционно считали потенциалнезависимой и нечувствительной к ТЕА утечкой. Ток утечки, активный в покое, стабилизирует мембранный потенциал ниже порога генерации потенциалов действия и ускоряет реполяризацию. Еще А. Ходжкин и А. Хаксли назвали эту структуру каналом утечки. С такими каналами связывают формирование потенциала покоя. Белковая структура, через которую осуществляется ток утечки, определена лишь несколько лет назад. Идентификация каналов, через которые осуществляется ток утечки, выявила, что это K+-каналы с двумя пороформирующими петлями в каждой субъединице и
четырьмя трансмембранными сегментами, как это показано на рис. 1-79 А. Две таких субъединицы формируют канал, названный K2P. Это K+-елективный канал с определенными параметрами проводимости, т.е. с потенциалнезависимым воротным механизмом, и предсказанным 50 лет назад выпрямлением. При физиологических условиях (высокая концентрация ионов K+ внутри и низкая снаружи) K2P проводит более значительный ток из клетки, чем внутрь клетки. Наблюдается поток ионов, направленный в большей степени наружу, чем внутрь клетки. Как ожидалось, для регуляторов возбудимости K2P-каналы находятся под сильным контролем множества химических и физических стимулов, включая напряжение кислорода в среде pH, липиды, механические растяжения, нейротрансмиттеры и рецепторы, связанные с G-белком. Каналы также служат молекулярными мишенями для некоторых летучих и местных анестетиков. Регуляция K2P-каналов связана с изменениями в их свойствах, например вероятности открытия каналов. Тем не менее некоторые регуляторные изменения, например фосфорилирование K2P, обеспечивают появление выпрямляющих свойств с чувствительностью к потенциалу.
На рисунке 1-79 Б представлена классификация K2P-каналов, причем часть из них, отмеченная на рисунке, проявляет механосенситивность. Наконец, на рис. 1-79 В показаны типичные вольтамперные характеристики, имеющие выпрямление. На нем представлено изменение вольтамперных характеристик в зависимости от величины рН для канала TASK-1, принадлежащего семье K2P-каналов.
Рис. 1-79. К+-каналы с двумя петлями в домене (two-P K+-channels - К2Р)
Потенциалы действия и токи, их формирующие
Рис. 1-80. Связь одиночного потенциала действия с основными ионными токами у разных клеток.
А - связь одиночного потенциала действия (А1) нервной клетки с ионными токами (А2). А1 - потенциал действия. Прямыми линиями помечены равновесный натриевый и калиевый потенциалы. А2 - две компоненты ионных токов, показанные через проводимость: входящий Na+-ток, осуществляемый через потенциалуправляемые Na+-каналы, и выходящий К+-ток, осуществляемый через потенциалуправляемые К+-каналы. На уровне критического потенциала наблюдается активация входящего Na+-тока, который инактивируется, когда мембранный потенциал достигнет положительных значений. Инактивация входящего Na+-тока происходит одновременно с активацией выходящего К+-тока. После достижения мембранного потенциала уровня потенциала покоя выходящий К+-ток инактивируется. Б - связь потенциала действия и самопроизвольного смещения мембранного потенциала до уровня критического потенциала с некоторыми ионными токами у нервной клетки с самопроизвольной ритмической активностью. Б1 - потенциал действия клетки с регулярной ритмической активностью. Б2 - компоненты ионных токов, формирующих этот потенциал действия. Показаны входящий Na+-ток (INa), создающий фазу деполяризации потенциала действия, выходящий К+-ток (IK), формирующий фазу реполяризации потенциала действия и быстрый временный выходящий К+-ток (Ito). Потенциал мембраны такой клетки определяется активностью 6 токов, которые были обнаружены у клеток с ритмической активностью. Во-первых, это, разумеется, ток, генерируемый электрогенным Na+, К+-насосом мембраны, и выходящий К+-ток утечки, создающие мембранный потенциал покоя. Кроме того, это входящий Na+-ток (или в отдельных случаях входящий - Са2+-ток), формирующий фазу деполяризации потенциала действия, и так называемый выходящий К+-ток (IK), формирующий фазу реполяризации потенциала действия. Наконец, это два типа ионных токов, смещающих мембранный потенциал до уровня критического потенциала в период между потенциалами действия. Первый тип - это сильный входящий Na+-ток, текущий по другим каналам, нежели вышеописанные и формирующие фазу деполяризации потенциала действия. Именно этот ионный ток смещает мембранный потенциал в сторону деполяризации к критическому потенциалу. К другому типу относится быстрый транзиторный выходящий К+-ток (fast transient K+-current - IA, или transient outward current - Ito), инактивация которого устраняется следовой гиперполяризацией и который активируется в промежутке между двумя потенциалами действия в подпороговой области мембранного потенциала. Кроме этих шести описанных ионных токов существуют еще два: медленный Na+-ток и медленный К+-ток. Эти медленные токи приводят к самоподдерживающимся осцилляциям мембранного потенциала, лежащим в основе периодических пачечных разрядов нейронов. В - связь потенциала действия клетки рабочего миокарда с ионными токами. В1 - форма потенциала действия рабочего кардиомиоцита. В2 - изменение во времени проводимости для деполяризующих (верхняя панель) или реполяризующих (нижняя панель) ионных токов во время потенциалов действия. Нарастание потенциала действия возникает, когда стимул выше порогового быстро деполяризует мембрану, активируя быстрые Na+-каналы, поэтому фаза быстрой деполяризации связана с входом Na+ в кардиомиоцит за счет резкого увеличения gNa. Входящий Na+-ток, осуществляемый через потенциалуправляемые Na+-каналы, не только очень быстро активируется, но и также быстро инактивируется. Инактивация Na+-каналов потенциалзависимая и происходит при достижении фазы деполяризации, достигает значений +25-30 мВ. Такая кинетика входящего Na+-тока определяет практически вертикальную форму фазы деполяризации потенциала действия. Фаза ранней или частичной реполяризации происходит за счет выхода K+ через ионные каналы мембраны, проводящие транзиторный выходящий ток (Ito). В результате такого транзиторного выхода положительно заряженных ионов клетка на короткое время частично реполяризуется. Эта частичная реполяризация активирует входящий Са2+-ток, и у потенциала действия начинается фаза плато. Г - связь потенциала действия узловой клетки сердца с ионными токами. В основе спонтанных изменений мембранного потенциала (Г1) в синоатриальном узле лежат три тока (Г2): неселективный входящий ток (If), который переносится катионами и не блокируется ТТХ, медленный входящий Са2+-ток (ICa) и выходящий К+-ток (IK)
Дата добавления: 2018-08-06; просмотров: 356; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!