Расчет равновесного потенциала для ионов 5 страница
На рисунке показан принцип фиксации тока, или current clamp (рис. 1-51 А2), а также принцип фиксации потенциала, или voltage clamp(рис. 1-51 Б2).
Применительно к рисунку (рис. 1-51 А2) липидный бислой можно уподобить конденсатору, две обкладки которого находятся на небольшом расстоянии друг от друга, и при подаче тока одна обкладка заряжается положительно, а другая - отрицательно.
Итак, если мы подаем на мембрану ток (рис. 1-51 А3), то его протекание через Rm описывается законом Ома:

где Vm - потенциал на Rm.
Протекание тока через емкость можно рассчитать следующим образом:

Таким образом, для общего тока, текущего в покое через мембрану, получаем:
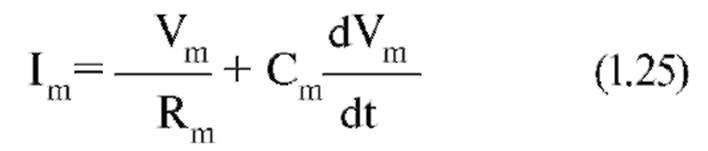
Емкостной компонент мембраны Сm обусловлен исключительно липидным бислоем, а резистивный компонент Rm - белками, образующими ионные каналы и встроенными в липидный бислой.
Если подать на мембрану прямоугольный импульс электрического тока, кривая изменения потенциала будет определяться RmxCm
Применительно к рис. 1-51 Б2 можно фиксировать потенциал и разрешить течь току.
Если внезапно менять напряжение от исходного значения до нового (в виде прямоугольного импульса; рис. 1-51 Б3), то быстро проходящий емкостной ток (Ic) течет по тому же принципу, как заряды текут на конденсатор. Ic максимален в начале прямоугольного импульса, когда заряд на конденсатор течет наиболее быстро, и затем уменьшается по экспоненте с постоянной времени RC. Если внезапно уменьшить напряжение до его исходного значения, Ic течет в противоположном направлении, по сравнению с регистрируемым в начале импульса. Таким образом, Ic появляется в виде коротких пиков в начале и в конце прямоугольного импульса. Изменение знака напряжения объясняется изменением направления тока в момент его прекращения, т.е. сначала происходит заряд, а затем разряд емкости, что связано с изменением на противоположное направление тока в цепи.
|
|
|
Итак, если Vm поддерживать постоянным, то Ic будет течь только очень короткое время, лишь в самый момент сдвига мембранного потенциала до нового значения. Затем этот ток прекратится, поскольку величина dVm/dt (скорость изменения мембранного потенциала) будет равна нулю. При этом мембранный ток становится простой функцией мембранной проводимости (gm) и напряжения (Vm):

Ток, который должен подаваться усилителем обратной связи для поддержания заданной величины мембранного потенциала, в точности равен общему мембранному току, протекающему при данном мембранном потенциале через участок мембраны, на котором напряжение фиксировано. Эти токи позволяют оценивать изменения общей ионной проводимости (а отсюда и специфических ионных проводимостей), вызываемые изменением мембранного потенциала.
|
|
|
В целом, метод был основан на том, что в известном выражении

емкостной ток становится равным нулю, если Vm задать равным константе. При этом Im становится равным Ii, т.е. ионный ток оказывается выведенным во внешнюю измерительную цепь.
Рис. 1-51. Сравнение методов регистрации с помощью обычного микроэлектрода (А) и с помощью patch- пипетки (Б)

Ионные токи
Результат применения метода фиксации потенциала представлен на рис. 1-52 А. На мембране поддерживается определенный потенциал (holding potential), равный, например, -60 мВ. Если с помощью прямоугольного импульса электрического тока сместить мембранный потенциал (деполяризовать мембрану) до 0 мВ, то в зарегистрированном токе можно различить три отдельные фазы. Вначале наблюдается так называемый емкостной ток, т.е. мгновенный пик тока (выделен зеленым цветом), направленного наружу, который обусловлен разрядом мембранной емкости. Поскольку она оказывается полностью разряженной, последующий ток определяют ионы, проходящие через ионные каналы. Вторая фаза представляет собой ток, направленный внутрь клетки (входящий ток Iin), т.е. вход катионов в цитозоль через ионные каналы мембраны. Эта фаза относительно кратковременна и переходит в третью фазу тока,
|
|
|
который течет из клетки (выходящий ток Iout) до тех пор, пока импульс электрического тока поддерживает деполяризацию мембраны.
Если мембрана резко деполяризована (рис. 1-52 Б), общий ионный ток (т.е. текущий после почти мгновенного разряда емкости) состоит из двух фаз - входящего и выходящего токов (синяя кривая). Поскольку ионный ток определяется ионами Na+ иК+ и зависит от их концентраций, то, изменяя концентрацию этих ионов, его можно разделить на компоненты. Было установлено, что если все ионы Na+, находящиеся во внешней среде, заменить на холин, который не проходит через мембрану, то входящий ток будет отсутствовать (коричневая кривая). Следовательно, он обусловлен ионами Na+. В этом случае выходящий ток приписывали ионам К+. Сначала ионы Na+ движутся по концентрационному градиенту, создавая входящий ток. Однако эта компонента быстро уменьшается и сменяется выходящим К+-током.
Рис. 1-52. Ионные токи, зарегистрированные методом фиксации потенциала.
А - ток, протекающий через мембрану (синяя кривая) при смещении потенциала до 0 мВ относительно поддерживаемого потенциала, равного -60 мВ (поддерживаемый и стимулирующий ток выделен красным цветом). Б - разделение мембранного тока (Im) на калиевую и натриевую компоненты: 1 - аксон находится в физиологическом растворе, I = INa + IK; 2 - натрий заменен на холин, I = IK; 3 - разность между 1 и 2, I = INa. Отклонение кривой вниз соответствует входящему току, а вверх соответствует выходящему току. Поддерживаемый потенциал мембраны клетки и его смещение обозначены красной кривой
|
|
|

Регистрация ионных токов методом фиксации потенциала
На рисунке 1-53 А представлен принцип регистрации тока при фиксации потенциала на клетке. Мембранный потенциал регистрируется микроэлектродом и подается на усилитель напряжения. С помощью электронной схемы с обратной связью его можно длительное время фиксировать (или поддерживать) на любом уровне путем пропускания тока через второй микроэлектрод, поэтому этот потенциал также называют поддерживаемым (holding potential). При помощи генератора прямоугольных импульсов электрического тока на мембране поддерживаемый потенциал можно смещать до некоторой новой величины и удерживать на этом уровне также с помощью электронной схемы с обратной связью. Ток, протекающий через этот участок мембраны при поддерживаемом потенциале или под влиянием приложенного напряжения, измеряют отдельным усилителем.
На рисунке 1-53 Б показаны примеры регистрации при типичных экспериментах с фиксацией потенциала у ооцита, в который экспрессированы потенциалуправляемые Na+-каналы. В этих экспериментах потенциал поддерживается на уровне -80 мВ. В первой части экспериментов тестирующая ступенька длиной 10 мс смещает потенциал от -80 до -120 мВ и вновь возвращает его к -80 мВ. В этом случае регистрируют только короткие емкостные пики в начале и в конце прямоугольного импульса. В промежутке между емкостными
пиками, т.е. на фоне самой ступеньки, не регистрируются никакие другие токи. Во второй части экспериментов тестирующая ступенька длиной 10 мс смещает потенциал от -80 мВ до -40 мВ и вновь возвращает его к -80 мВ. В этом случае в промежутке между емкостными пиками, т.е. на фоне самой ступеньки, регистрируется входящий ток. Этот ионный ток, протекающий через экспрессированные в ооцит потенциалуправляемые №+-каналы. Часто этот ток называют макроскопическим, поскольку он протекает через большую популяцию каналов в целой клетке.
Для понимания механизма удобно рассмотреть электрическую модель аксона. На рисунке 1-53 В сопротивление r представляет собой сопротивление осевого электрода, связывающего разные участки мембраны. Предположим, что моделируемый аксон находится в большом объеме раствора, так что наружная жидкость эквипотенциальна и на схеме может быть представлена проводником без сопротивления. На рисунке показан один элемент мембраны, а следует представить себе большое число аналогичных элементов, связанных между собой и образующих непрерывный кабель. Каждый элемент содержит емкость мембраны С, калиевую и натриевую батареи Ек и ЕNa и сопротивления RK и RNa. Сопротивление утечки Rl и батарея Еl введены для учета движения ионов, проходящих по каналам, которые не изменяются во время активности. Однако ток утечки мал, и в первом приближении его можно не принимать во внимание.
Рис. 1-53. Двухэлектродный метод фиксации потенциала.
А - схема опыта. Б - регистрируемые токи на фоне гиперполяризации и деполяризации. В - эквивалентная электрическая схема элемента возбудимой мембраны аксона. Обозначения: r - сопротивление осевого электрода, введенного в аксон; С - емкость мембраны; VK - калиевая батарея; RK - сопротивление К+-канала; VNa - натриевая батарея; RNa - сопротивление Nа+-канала; Rl - сопротивление утечки; Vl - батарея утечки

Na+- и К+- проводимость
Зная величину калиевого и натриевого тока, нетрудно оценить проводимость мембраны для каждого иона. При подпороговых значениях деполяризации выходящий калиевый ток больше входящего натриевого, что препятствует достижению мембранным потенциалом порога.
Как только входящий Na+-ток хотя бы слегка превысит выходящий К+-ток, мембранный потенциал неизбежно начнет смещаться регенеративным образом в сторону ENa .
Зная величину калиевого и натриевого тока, можно оценить проводимость мембраны для каждого иона. На рисунке 1-54 А показаны изменения проводимости, вызванные быстрым смещением потенциала внутри волокна до 0 мВ относительно поддерживаемого потенциала, равного -56 мВ. Смещение мембранного потенциала до нуля эквивалентно короткому замыканию мембраны. В этом случае конденсатор С сразу разряжается, и с этого момента ток создают только ионы, проходящие по каналам, имеющим сопротивление RNa иRK.
Натриевая проводимость (gNa) в ответ на смещение мембранного потенциала относительно поддерживаемого потенциала изменяется от крайне низкой величины и быстро нарастает, а затем экспоненциально уменьшается. Это экспоненциальное уменьшение gNa может быть быстрым или медленным, что определяется длительностью ступеньки деполяризации. Если смещение потенциала относительно поддерживаемого потенциала (например, потенциала покоя) было кратковременным (как это показано красной пунктирной линией; рис. 1-54 А, красная кривая, импульс 1), и потенциал покоя был быстро восстановлен, gNa быстро возвращается к состоянию покоя (пунктирная красная кривая). Если сразу же подать второй аналогичный импульс, вызывающий смещение потенциала относительно поддерживаемого потенциала, то это также вызывает аналогичное увеличение gNa. Если же деполяризация продолжительна (рис. 1-54 А, красная кривая, импульс 2 или 3), gNa быстро увеличивается, но уменьшается более медленно (рис. 1-54 А, сплошная кривая)
вследствие процесса, называемого инактивацией. В этом случае, чтобы второй импульс после инактивации натриевых каналов мог снова вызвать изменение проницаемости для натрия, мембрана в течение нескольких миллисекунд до нанесения этого импульса должна быть реполяризована. Что же касается калиевой проводимости (gK), то во всяком случае в аксоне кальмара не обнаружено инактивации, и высокая gK сохраняется все время, пока мембрана деполяризована. Так, если смещение потенциала относительно поддерживаемого потенциала было не очень длительным (рис. 1-54 А, красная кривая, импульс 2), высокая gK сохраняется до возвращения потенциала к поддерживаемому потенциалу (рис. 1-54 А, пунктирная коричневая кривая). Если же смещение потенциала относительно поддерживаемого потенциала было длительным (рис. 1-54 А, красная кривая, импульс 3), высокая gK сохраняется все время, пока мембрана деполяризована (рис. 1-54 А, сплошная коричневая кривая).
Калиевая проводимость (gK) нарастает от небольшой, но вполне определенной величины. Это изменение начинается не сразу. Кривая увеличения gK имеет S-образную форму, и проводимость выходит на постоянный уровень через 5-6 мс. Задержанный сдвиг gKдостигает максимума, и это начинает возвращать мембранный потенциал к уровню потенциала покоя (а часто сдвигает его и несколько дальше этого уровня). Кроме того, приходит в действие другой независимый механизм: деполяризация мембраны вызывает также (с задержкой) инактивацию gNa, что тоже способствует возвращению мембранного потенциала к уровню потенциала покоя.
Таким образом, деполяризация гигантского аксона кальмара запускает три процесса:
быстрое нарастание gNa;
• задержанное нарастание калиевой проводимости, которая не инактивируется (но выключается при реполяризации мембраны);
• задержанную инактивацию gNa (рис. 1-54 А, Б). Рассчитанные значения потенциала действия
(А.Л. Ходжкин и А.Ф. Хаксли) были похожи на зарегистрированный потенциал действия (рис. 1-54 В).
Рис. 1-54. Натриевая и калиевая проводимости и теоретическая реконструкция потенциала действия.
А - изменения во времени gNa и gK при деполяризации на 56 мВ, т.е. от поддерживаемого потенциала, равного -56 мВ, до 0 мВ. Поддерживаемый потенциал мембраны клетки и его смещение обозначены красной кривой, gNa и gK - бордовой и коричневыми кривыми соответственно. Сплошными линиями обозначены продолжительная деполяризация и изменения gNa и gK в этом случае; пунктирными линиями показаны изменения gNa и gK в ответ на более короткие ступени деполяризации gNa и gK. Б - рассчитанные изменения gNa и gKпри развитии потенциала действия (V). В - сравнение рассчитанного потенциала действия (В1) с реальным потенциалом действия, зарегистрированным в гигантском аксоне кальмара (В2). Рассчитанная скорость проведения потенциала действия составляла 18,8 м/с, а полученная в эксперименте - 21,2 м/с

Токи при разных ступеньках потенциала
Рассмотрим ответ мембраны клетки на ступенчатое смещение мембранного потенциала относительно поддерживаемого потенциала (потенциала покоя). Пусть мембранный потенциал поддерживается на величине -60 мВ, равной величине потенциала покоя клетки. В этом случае мы не зарегистрируем ни входящего, ни выходящего тока, и прибор, регистрирующий ток, будет показывать нулевую линию. Теперь ступенчато сместим поддерживаемый потенциал на 20 мВ в сторону деполяризации (рис. 1-55 А - верхний фрагмент). На приборе, регистрирующем текущий через мембрану ток, можно будет увидеть осцилляцию нулевой линии, включающую отклонение тока в отрицательное направление (т.е. входящий ток) и последующее отклонение в положительном (т.е. выходящий ток) (рис. 1-55 Б - верхний фрагмент). Однако и входящий, и выходящий токи при подобном смещении мембранного потенциала малы. Теперь ступенчато сместим поддерживаемый потенциал на 40 мВ в сторону деполяризации. На приборе можно будет видеть увеличение амплитуд входящего и выходящего токов. Сместим поддерживаемый потенциал на 60 мВ, т.е. до 0 мВ. На приборе мы увидим максимальное увеличение амплитуды входящего и выходящего токов. При более высоких величинах деполяризации входящий ток становится меньше и при смещении мембранного потенциала от 110 до 120 мВ, сдвигающей потенциал мембраны в зону от +50 до +60 мВ, обращается в нуль (до потенциала реверсии). Если смещение потенциала будет еще больше (на 160 мВ), входящий ток изменит знак и будет течь наружу. Выходящий ток с увеличением ступенек ведет себя противоположным образом: по мере того как мембрана все больше деполяризуется,
он увеличивается. Выходящий ток уменьшается только тогда, когда мембранный потенциал изменяется в сторону гиперполяризации (после того как вначале он был быстро сдвинут в сторону деполяризации ступенькой порядка 84 мВ). Потенциал реверсии для выходящего тока равен около -80 мВ. Изменения входящего и выходящего токов в зависимости от мембранного потенциала легко представить в виде вольтамперных характеристик (рис. 1-55 В). На этих графиках пики входящего тока и установившиеся значения выходящего откладываются по оси абсцисс как функции смещения мембранного потенциала относительно поддерживаемого потенциала. Как видно из таких графиков, деполяризующие ступеньки активируют как входящий, так и выходящий токи. Вначале с увеличением деполяризации оба тока увеличиваются по амплитуде. Затем входящий ток уменьшается и при деполяризующих ступеньках более 110 мВ меняет знак.
Входящий ток объясняется тем, что в результате увеличения gNa, вызываемого деполяризацией, ионы Na+ устремляются внутрь клетки. Исходя из увеличения gNa, этот ток прекращается при +55 мВ - потенциале равновесия Нернста для Na+ (ЕNa). При ЕNa сила, обусловленная электрическим потенциалом, равна по величине и противоположна по направлению силе, создаваемой градиентом концентрации, так что суммарного натриевого тока нет. При еще большей деполяризации первая из этих сил становится больше второй, и ионы Na+ начинают выходить из клетки наружу, тем самым изменяя направление тока. Подобным образом особенности выходящего тока определяются тем, что его носителями служат ионы К+. По мере деполяризации мембраны силы электрического поля, стремящиеся удержать К+ внутри клетки, уменьшаются, приводя ко все большему преобладанию направленных наружу сил, обусловленных градиентом концентрации К+.
Рис. 1-55. Фиксация потенциала на гигантском аксоне кальмара.
Дата добавления: 2018-08-06; просмотров: 339; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!