Расчет равновесного потенциала для ионов 2 страница

Таким образом, измерение τm важно также для оценки Cm. Пусть τm равно 100 мс, a Rm - 100 000 Омхсм2, тогда емкость Cm будет равна 1 мФ/см2:

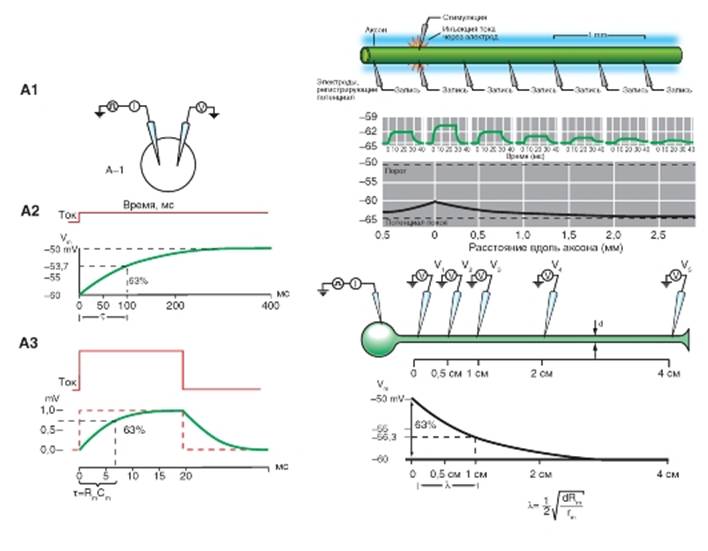
Постоянная длины мембраны
Постоянная длины мембраны (λ) - это расстояние от точки в нейроне, где при помощи внутриклеточного электрода был изменен мембранный потенциал, до той точки на аксоне, где этот потенциал потеряет 63% или иначе 1 = 1/e своей первоначальной величины.
Изменения потенциала в различных точках аксона можно измерить с помощью введения регистрирующих электродов (V1, V2, V3, V4) на различных расстояниях вдоль аксона (рис. 1-38 Б1). Пусть аксон имеет большой диаметр, сравнимый с размером клеточного тела. Тогда постоянная длины (в сантиметрах) будет определяться формулой:
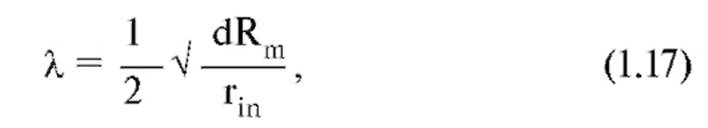
где Rm - удельное сопротивление мембраны; rin - удельное сопротивление аксоплазмы; d - диаметр аксона. Мы приводим лишь конечную формулу, поскольку в основе ее вывода лежат те же принципы, что и для постоянной времени.
Постоянная длины мембраны - это расстояние, на которое вдоль аксона электротонически распространяются подпороговые сигналы. Если нейрон имеет сопротивление Rm= 100 000 Омхсм2 и rin= 50 Омхсм2, постоянная длины при диаметре аксона 20 мкм равна 1 см. Но для аксонов малого диаметра d постоянная длины может быть значительно меньше и достигать 0,1 см.
Рис. 1-38. Определение постоянных времени и длины.
А - постоянная времени. На панели А1 изображена схема эксперимента. Через стимулирующий внутриклеточный микроэлектрод на клетку подается импульс электрического тока, а изменения мембранного потенциала измеряют с помощью регистрирующего внутриклеточного микроэлектрода. На панели А2 показано изменение мембранного потенциала во времени (нижняя кривая) на фоне подачи длительного импульса постоянного электрического тока (верхняя кривая). Ток вызывает деполяризацию величиной 10 мВ и сдвигает мембранный потенциал с -60 до -50 мВ. Время, необходимое для достижения 63,2% этого изменения, называют постоянной времени клеточной мембраны. В данном примере она равна 100 мс. На панели А3 показано изменение мембранного потенциала во времени (нижняя кривая) на фоне подачи короткого импульса постоянного электрического тока (верхняя кривая). Б - постоянная длины. На панели Б1 изображен принцип эксперимента. Через стимулирующий внутриклеточный микроэлектрод, введенный в аксон, подается импульс электрического тока, который распространяется электротонически по аксону, а изменения мембранного потенциала измеряются внутриклеточными регистрирующими микроэлектродами в точке стимуляции (V1) и в различных точках вдоль аксона (V2, V3, V4, V5). На панели Б2 показаны изменения мембранного потенциала в зависимости от расстояния (сверху) и график уменьшения амплитуд. На панели Б3 изображена схема эксперимента. Через стимулирующий внутриклеточный микроэлектрод, введенный в тело нейрона, подается импульс электрического тока, который распространяется электротонически по аксону, а изменения мембранного потенциала измеряются внутриклеточными регистрирующими микроэлектродами у основания аксона (V1) и в различных точках вдоль него (V2, V3, V4, V5). Внизу показаны изменения мембранного потенциала в зависимости от расстояния. В теле клетки ток вызывает деполяризацию величиной 10 мВ и сдвигает мембранный потенциал с -60 до -50 мВ (V1). На расстоянии 1 см (V2) это изменение потенциала уменьшается на 63%, достигая 37% своего исходного значения (3,7 мВ). Это расстояние (1 см) называют постоянной длины мембраны аксона
|
|
|
|
|
|
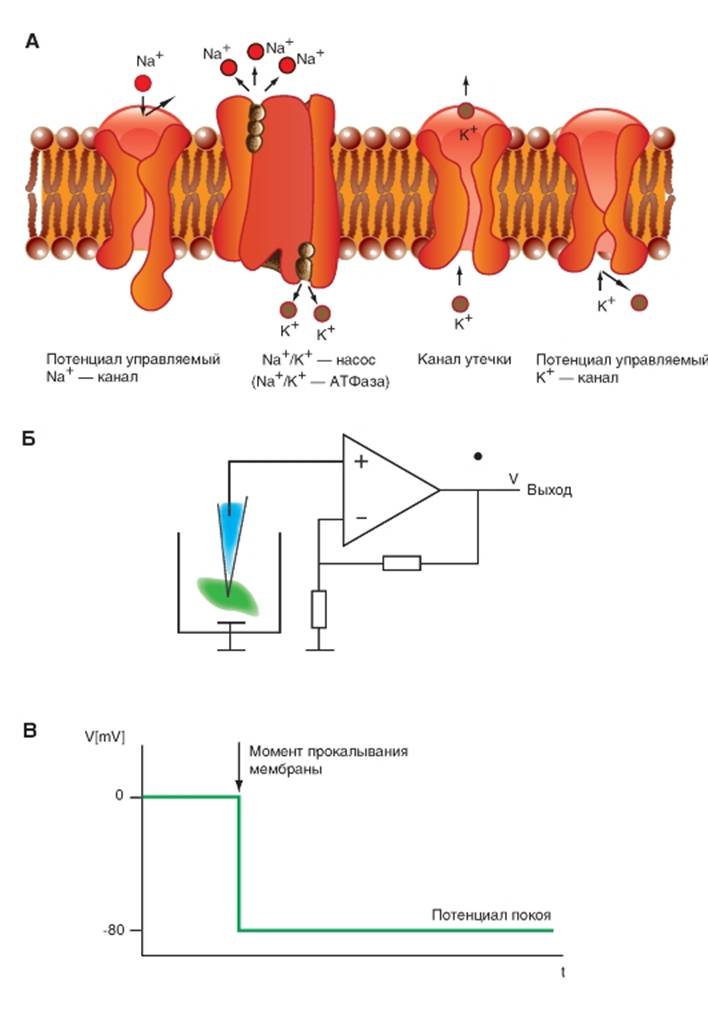
Потенциал покоя и его механизмы
Потенциал покоя клетки определяется работой Na+/K+-АТФазы и выходом ионов K+ по каналам утечки при очень незначительном входе ионов Na+ через них (рис. 1-39 А). Распределение ионов относительно мембраны, а именно внутри клетки, поддерживается механизмом активного транспорта Na+ и K+, или электрогенным Na+/K+-насосом, который удерживает внутри клетки концентрацию Na+ на низком, а концентрацию K+ на высоком уровнях, транспортируя Na+ из клетки наружу, а K+ из наружной среды внутрь. Поскольку равновесный потенциал для ионов Na+ равен +55 мВ, а потенциал мембраны клетки лежит в диапазоне от -60 до -80 мВ, существует движущая сила для ионов Na+, и они устремляются по каналам утечки вовнутрь клетки, что могло бы деполяризовать ее мембрану. Однако каналы утечки проницаемы и для ионов K+, для которых сохраняется электрохимический градиент, направленный из внутриклеточной области к внеклеточной среде. Так как равновесный потенциал ионов K+ равен примерно -90 мВ, через мембрану одновременно проходят и ионы K+ тем самым, противодействуя деполяризации, обусловленной входящим током
|
|
|
ионов Na+. При этом работа Na+/K+-АТФазы удерживает внутри клетки концентрацию Na+ на низком, а концентрацию K+ на высоком уровнях. Имеющуюся в клетке трансмембранную разность потенциалов можно зарегистрировать.
Если микроэлектрод, подключенный к предварительному усилителю, показанному на рис. 1-39 Б, подключить к измерительной схеме и ввести в физиологичский раствор, где расположены ткань или клетки, при этом замкнуть цепь через индифферентный электрод, то при сбалансированном усилителе и компенсации физико-химических свойств микроэлектрода на экране осциллографа мы будем регистрировать линию (рис. 1-39 В), соответствующую измерительному нулю, равному нулю при закороченных входах усилителя. Теперь если подвести при помощи микроманипулятора микроэлектрод вплотную к мембране клетки и проколоть ее, то на экране осциллографа можно зарегистрировать падение нулевого потенциала и разность потенциалов между внутренней и наружной средами клетки, величина которой для разных клеток лежит в диапазоне от -40 до -80 мВ. Именно эта разность, представленная на рис. 1-39 В, служит потенциалом покоя клетки.
|
|
|
Рис. 1-39. Потенциал покоя клетки и принцип его регистрации.
А - структуры мембраны, формирующие потенциал покоя. На панели А представлена мембрана клетки, канал утечки, Nа+-канал, К+-канал и -АТФаза, которая выкачивает ионы Na+ из клетки против его электрохимического градиента, одновременно вводя ионы K+ в клетку также против его электрохимического градиента. Каналы утечки, через которые осуществляется очень незначительный вход ионов Na+ в клетку и превышающий его в 100 раз выход ионов К+ из клетки. За счет каналов утечки и работы Na+/K+-АТФазы возникает негативный внутриклеточный потенциал мембраны клетки - потенциала покоя. Б - измерительная схема. В - потенциал покоя
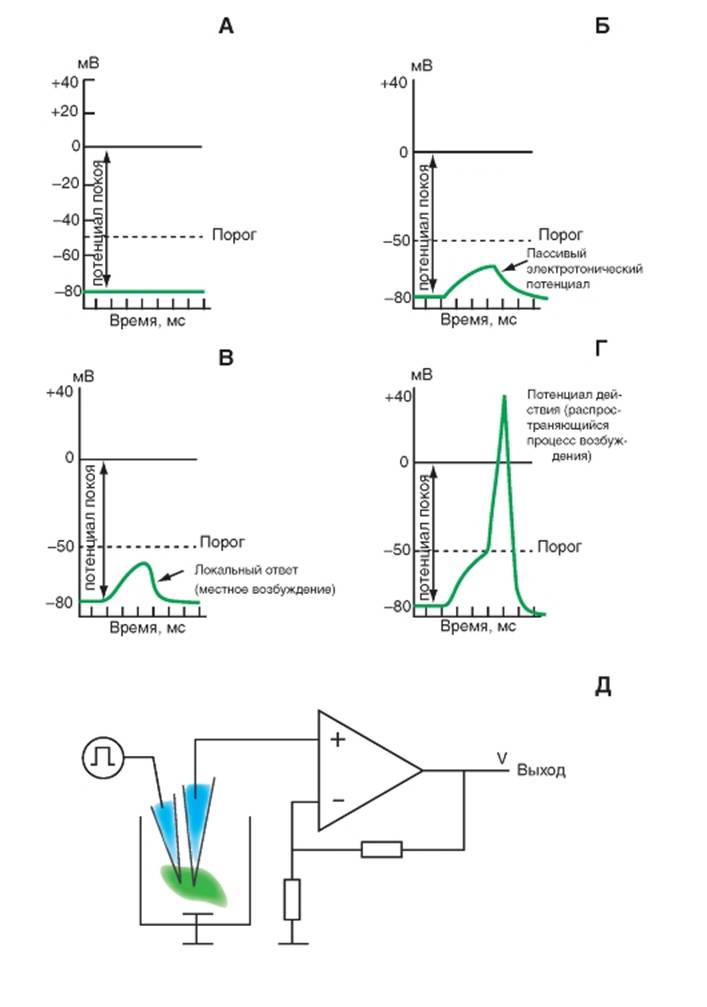
Потенциалы клетки, возникающие при деполяризации ее мембраны, и принцип искусственного смещения потенциала
Помимо потенциала покоя (рис. 1-40 А), пассивный транспорт ионов через ионные каналы мембраны определяет возникновение еще трех потенциалов, представленных на рисунке. К ним принадлежат пассивный электротонический потенциал (рис. 1-40 Б), локальный ответ (рис. 1-40 В) и потенциал действия (рис. 1-40 Г). Однако для их возникновения требуется поляризация мембраны клетки. Эта поляризация может осуществляться как внеклеточно, что обычно используют на нервных волокнах, так и внутриклеточно, что применяют на клетках. Пассивный электротонический потенциал зарождается при заведомо подпороговом смещении потенциала покоя. Локальный ответ возникает при подпороговом, но близком к порогу смещении потенциала покоя. Наконец, потенциал действия зарождается, когда смещение потенциала покоя доведено до пороговой величины.
Однако, чтобы зарегистрировать эти потенциалы, необходимо искусственное смещение потенциала покоя клетки. Возможность внутриклеточной поляризации мембраны, включающей деполяризацию и гиперполяризацию, крайне необходима
для изучения свойств мембраны. Искусственное смещение мембранного потенциала дает возможность не только изучать пассивные электрические характеристики клеток, но и исследовать межклеточные взаимодействия, осуществляемые непосредственно через высокопроницаемые контакты мембран клеток. Однако возможность внутриклеточной поляризации мембран должна непременно сочетаться с возможностью одновременной регистрации биоэлектрической активности клеток и тех изменений этой активности, которые возникают вследствие приложения электрического тока.
В наиболее примитивном виде принцип внутриклеточной поляризации мембран при возможности одновременной регистрации биоэлектрической активности клеток представлен на рис. 1-40 Д. В этом случае в клетку одновременно вводят два микроэлектрода, один из которых поляризующий, а другой - регистрирующий. На экране двухлучевого осциллографа можно видеть весь процесс: на первом канале регистрируется величина поляризующего сигнала, а на втором канале - мембранный потенциал и его искусственное смещение, вызванное поляризацией мембраны. Следует помнить, что прямое подключение поляризующего микроэлектрода к генератору невозможно, для этого существуют специальные схемы.
Рис. 1-40. Изменение мембранного потенциала в зависимости от силы раздражения.
А - потенциал покоя. Б - пассивный электротонический потенциал. В - локальный ответ. Г - потенциал действия. Д - выход
Пассивный электротонический потенциал
Пассивный электротонический потенциалможно зарегистрировать с помощью одного внутриклеточного микроэлектрода, если через второй микроэлектрод внутриклеточно поляризовать клетку прямоугольным импульсом электрического тока, не достигающим критического потенциала (подпороговым) (рис. 1-41 А).
Пассивный электротонический потенциал возникает в ответ на подпороговый импульс электрического тока, который не приводит к открытию потенциалуправляемых ионных каналов и определяется только емкостными и резистивными свойствами мембраны клетки. Емкостной компонент мембраны обусловлен исключительно липидным бислоем, а резистивный компонент - белками, образующими ионные каналы и встроенными в липидный бислой.
Липидный бислой можно уподобить конденсатору, две обкладки которого находятся на небольшом расстоянии друг от друга, и при подаче тока одна обкладка заряжается положительно, а другая - отрицательно. Ионные каналы липидного бислоя или в этом случае мембраны клетки обладают проводимостью, и, следовательно, мембрана характеризуется электрическим сопротивлением.
На рисунке 1-41 Б изображена эквивалентная электрическая схема мембраны клетки, а на рис. 1-41 В показана эквивалентная электрическая схема фрагмента мембраны. На подаваемый ток и изменения сопротивления и напряжения эта схема реагирует точно так же, как и сама мембрана.
Данная схема включает емкостной компонент мембраны (Сm) и резистивный компонент (Rm), а также учитывает сопротивления внешней среды rout и сопротивление цитозоля rin. Поскольку значения rout и rin малы, ими можно пренебречь, хотя это достаточно грубая аппроксимация. Кроме того, rout много меньше rin
Если на мембрану через один микроэлектрод подать подпороговый прямоугольный импульс электрического тока положительной полярности (рис. 1-41 Г - импульс красного цвета), то усилитель, подключенный ко второму микроэлектроду, зарегистрирует изменения мембранного потенциала, форма которого отличается от прямоугольника (зеленая кривая). Это и есть пассивный электротонический потенциал.
Если мы подаем на мембрану ток, то его протекание через Rm описывается законом Ома:

где Vm - потенциал на Rm.
Протекание тока через емкость можно рассчитать следующим образом:

Таким образом, для общего тока, текущего в покое через мембрану, получаем:
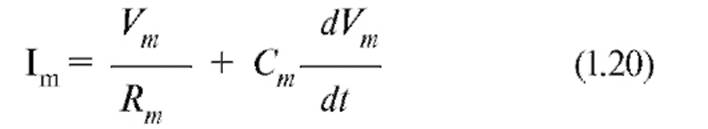
Емкостной компонент мембраны Сm обусловлен исключительно липидным бислоем, а резистивный компонент Rm - белками, образующими ионные каналы и встроенными в липидный бислой. Интегрируя выражение, получим:
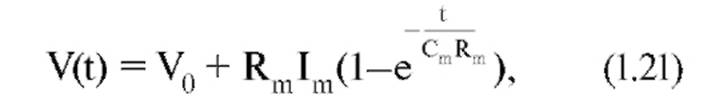
где Rm Сm =τm , что называется постоянной време- ни(временем релаксации мембраны). Постоянная времени мембраны τ - одна из важнейших характеристик пассивного электротонического потенциала, которая отражает пассивные свойства мембраны. Последнее уравнение крайне важно, поскольку величины τm и Rm можно, в отличие от Cm, зарегистрировать экспериментально и, следовательно, таким образом рассчитать Cm. С помощью постоянной времени и Rm можно полностью описать пассивные свойства эквивалентной схемы (или мембраны).
Постоянная времени характеризует временной ход изменений мембранного потенциала, т.е. скорость, с которой он меняется при переходе от одного значения к другому. Постоянная времени мембраны - это время, необходимое для того, чтобы импульс постоянного тока зарядил емкость мембраны на 63% (см. рис. 1-41 Г).
Сходная картина возникает, если на мембрану через один микроэлектрод подать аналогичный по силе прямоугольный импульс электрического тока отрицательной полярности (рис. 1-41 Д - импульс красного цвета). В этом случае пассивный электротонический потенциал будет направлен в отрицательную область (зеленая кривая).
Рис. 1-41. Пассивный электротонический потенциал

Механизм пассивного электротонического потенциала
Пассивный электротонический потенциал определяется только емкостными и резистивными свойствами мембраны клетки, но мы относим его к потенциалам клетки, связанным с пассивным ионным транспортом, т.е. транспортом через ионные каналы. Это обусловлено тем, что электрическая цепь, включающая положительный вход усилителя, микроэлектрод, клетку, индифферентный электрод и отрицательный вход усилителя, может быть замкнута только в том случае, если мембрана имеет возможность проводить через себя электрический ток, носителем которого в системе клетки и внеклеточного раствора служат ионы. Эту возможность дают постоянно открытые каналы утечки.Однако пассивный электротонический потенциал не приводит к открытию потенциалуправляемых ионных каналов. В том случае емкостные и резистивные свойства мембраны клетки достаточно постоянны. Емкость в основном определяется липидным бислоем, а сопротивление клетки зависит от сопротивления, которое определяется открытыми каналами утечки.
На рисунке 1-42 А показан пассивный электротонический потенциал, в котором мы искусственно выделяем отдельные фазы (0 - потенциал покоя, 1 - фаза нарастания, 2 - фаза падения), а также клетка в каждую из этих фаз. Из рисунка следует, что на фоне потенциала покоя, т.е. в фазу 0, открыты только каналы утечки, через которые ионы К+ выходят из клетки, а ионы Na+ в незначительном количестве входят в клетку. Это соотношение обычно принято как К+: Na+=100:1. При этом потенциалуправляемые Na+- каналы, обеспечивающие входящий ток, и потенциалуправляемые К+-каналы, обеспечивающие выходящий ток, закрыты (находятся в состоянии покоя). В фазу 1 и фазу 2 также открыты только каналы утечки, через которые ионы К+ выходят из клетки, а ионы Na+ в незначительном количестве входят в клетку. Соотношение K+:Na+=100:1 не меняется. При этом потенциалуправляемые Na+-каналы, обеспечивающие входящий ток, и потенциалуправляемые К+-каналы, обеспечивающие выходящий ток, также закрыты (находятся в состоянии покоя). При нарастании потенциала (фаза 1), как и при его падении (фаза 2) до уровня
потенциала покоя, не происходит никаких изменений ионных токов, поскольку, как отмечалось, пассивный электротонический потенциал определяется только емкостными и резистивными свойствами мембраны.
Характерная особенность пассивного электротонического потенциала состоит в равенстве скоростей нарастания и спада экспоненты.
Для различных клеток значения τm варьируют от одной до нескольких сотен миллисекунд, однако для одной и той же клетки эта величина равна вне зависимости от длительности и направления поляризующего импульса электрического тока. Это еще одна особенность пассивного электротонического потенциала.
Наконец, если дискретно увеличивать амплитуду поляризующего импульса электрического тока, оставаясь тем не менее в подпороговом диапазоне его силы (рис. 1-42 В1), то амплитуда каждого пассивного электротонического потенциала будет равна амплитуде импульса тока, его вызывающего. В этом проявляется строго линейная зависимость между амплитудами электрического стимула и пассивного электротонического потенциала (рис. 1-42 В2). Значительное увеличение длительности поляризующего импульса не меняет этой закономерности (рис. 1-42 Г1, Г2). Ранее мы обсуждали, что как только емкость зарядится до потенциала, равного подаваемому на нее, емкостной ток прекратится. Таким образом, она не препятствует изменениям потенциала, а только замедляет его нарастание и падение.
Наклеткуможноподатьпрямоугольныйимпульс электрического тока сверхпороговой амплитуды, т.е. величиной в несколько вольт или даже в несколько десятков вольт, без ущерба для данной клетки, если длительность этого импульса будет меньше значения τm для данной клетки. В этом случае емкость мембраны не успевает полностью зарядиться и, следовательно, клетка не набирает столь большой потенциал (рис. 1-42 Д1, Д2).
Наконец, последняя особенность пассивного электротонического потенциала заключается в том, что в волокне, например аксоне, он распространяется с затуханием, характеризующим постоянную длину мембраны λ, т.е. расстояние вдоль аксона, на котором напряжение, приложенное в одной точке нейрона, потеряет 63% (1-1/e) своей первоначальной величины.
Рис. 1-42. Влияние различных прямоугольных импульсов электрического тока на форму пассивного электротонического потенциала
Дата добавления: 2018-08-06; просмотров: 773; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!