Образовавшийся ацил-КоА является активной формой жирной кислоты.Синтез триацилглицеролов
Образовавшиеся глицерол-3-фосфат и ацил-КоА жирных кислот – исходные соединения для биосинтеза жира (см. рис. 11, Этап V).
На первом этапе синтеза триацилглицеролов образуется 3-фосфатидная кислота (диацилглицерол-3-фосфат), при этом ацильные остатки КоА-производных жирных кислот переносятся специфическими ацил-трансферазами на спиртовые группы глицерол-3-фосфата:
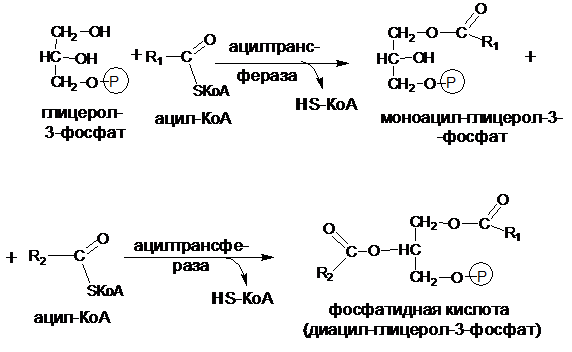
На втором этапе фосфатидная кислота гидролизуется фосфатидатфосфатазой с образованием 1,2-диацилглицерола:

Затем 1,2-диацилглицерол превращается в триацилглицерол путем переноса остатка КоА-производного третьей жирной кислоты. Этот перенос катализируется ферментом диацилглицеролацилтрансферазой:

42.
Распад (катаболизм) фосфолипидов
Фосфолипиды также как и жиры, активно распадаются в тканях животных и растений.
Гидролиз фосфолипидов осуществляют несколько фосфолипаз, различающихся по расщеплению определенных связей и получивших название фосфолипаз А, С и Д:
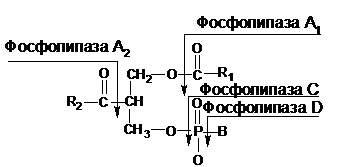
Здесь R1 и R2 – углеводородные радикалы; В – остатки азотсодержащих одноатомных спиртов (холина, этаноламина, серина).
Фосфолипаза А1 отщепляет жирные кислоты по α-положению; фосфолипаза А2 осуществляет ту же самую реакцию по β-положению; фосфолипаза С отщепляет фосфорилированные азотсодержащие спирты от фосфолипидов; фосфолипаза D гидролизует фосфолипиды с образованием азотсодержащего спирта и фосфатидной кислоты.
|
|
|
В растениях обнаружены все виды расщепления. Фосфолипазы А обнаружены также у животных и микроорганизмов. Фосфолипазы локализованы преимущественно в лизосомах. Конечными продуктами гидролиза фосфолипидов является глицерин, жирные кислоты, азотистые спирты и фосфорная кислота.
Синтез фосфолипидов
В отличие от триацилглицеролов и жирных кислот фосфолипиды не являются существенным энергетическим материалом. Фосфолипиды, как было сказано ранее, играют важную роль в структуре и функции клеточных мембран.
Биосинтез фосфолипидов интенсивно происходит в печени, стенке кишечника, молочной железе и других тканях. Наиболее важные фосфолипиды синтезируются главным образом в эндоплазматической сетке клетки.
Центральную роль в биосинтезе фосфолипидов играют L-диацилглицеролы (в синтезе фосфатидилхолинов и фосфатидилэтаноламинов), фосфатидная кислота (в синтезе фосфатидилинозитов) и сфингозин (в синтезе сфингомиелинов).
В этих биосинтезах принимает участие цитидинтрифосфат (ЦТФ), выполняющий роль активатора.
Основными компонентами мембран являются фосфатидилэтаноламин и фосфатидилхолин.
В процессе синтеза фосфатидилэтаноламина вначале происходит активация этаноамина под действием этаноламинкиназы:
|
|
|

Этаноламинфосфат реагирует затем с цитидинтрифосфатом (ЦТФ), в результате чего образуется цитидиндифосфатэтаноламин (ЦДФ-этаноламин) и пирофосфат (РРі). Эту реакцию катализирует фосфоэтаноламинцитидинтрансфераза:
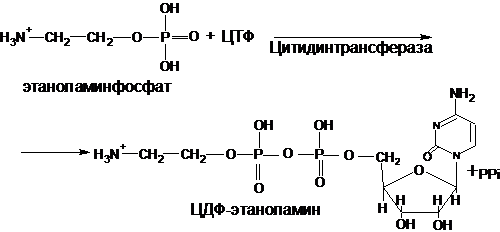
Активированный ЦДФ–этаноламин при участии фермента этаноламинфосфаттрансферазы вступает в реакцию с L–диацилглицеролом с образованием фосфатидиламина:

L-диацилглицерол образуется при гидролизе фосфатидной кислоты. Аналогичные реакции ведут к образованию ЦДФ-холина.; последний вступает в реакцию с L-диацилглицеролом, в результате чего образуется фосфатидилхолин.
В синтезе фосфатидилэтаноламина и фосфатидилхолина важную роль играет ЦТФ. Подобно тому синтезу олиго- и полисахаридов, мы и здесь видим, каким образом нуклеотиды могут выполнять функцию переносчиков опреденных химических групп в обмене веществ клеток.
Помимо синтеза фосфолипидов таким способом, в клетке они легко подвергаются взаимопревращениям. Фосфатидилэтаноламин является предшественником фосфатидилхолина. Так, в печени в результате последовательного переноса трех метильных групп от трех молекул S-аденозилметионина образуется фосфатидилхолин:
|
|
|

В переносе метильных групп важную роль играют тетрогидрофолевая кислота (витамин Вс) и метилкобаломин ( витамин В12).
Синтез фосфатидилсерина происходит другим путем с предварительным вовлечением фосфатидной кислоты:
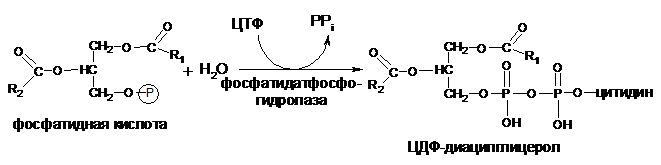
Затем происходит перенос серина на фосфатидильный остаток с образованием фосфатидилсерина:
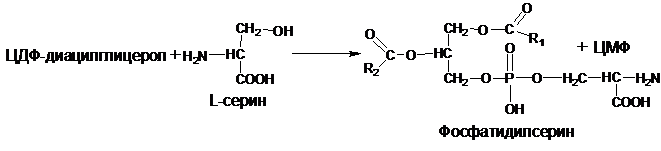
У млекопитающих фосфатидилсерин образуется в реакции обмена этаноламина на серин следующим путем:
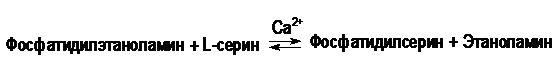
Таким же путем образуется фосфатидилинозитол.
В биосинтезе сфингомиелина принимает участие церамид (N-ацетилсфингозин), который образуется при взаимодействии спирта сфингозина и ацетил-КоА. Сфингомиелин синтезируется в результате взаимодействия церамида с ЦДФ-холином:
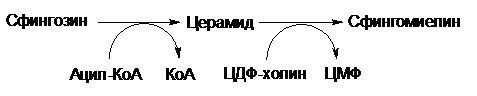
Все выше синтезированные фосфолипиды поступают с помощью липидпереносящих белков из цитоплазмы к мембранам и участвуют в их построении.
43.Взаимосвязь белкового и липидного обменов
Рассмотренные взаимосвязи «белки Û углеводы» и «углеводы Û липиды» дают основание для объединения их в единую цепь «белки Û углеводы Û липиды», в которой углеводы являются связующим звеном между белками и липидами.
|
|
|
Один из основных продуктов расщепления липидов – ацетил-КоА, включаясь в ЦТК, образует кетокислоты, аминирование которых дает аминокислоты.Из другого продукта гидролиза липидов глицерина – в результате цепи превращений синтезируются циклические аминокислоты.В известной мере возможен и обратный процесс синтеза липидов за счет распадающихся белков. Продукты дезаминирования аминокислот через ЦТК и другие метаболические процессы образуют ПВК, при окислительном декарбоксилировании которого возникает ацетил-КоА – исходное соединение для синтеза жирных кислот и других компонентов липидов.Из всего вышесказанного видно, что превращения веществ в организме тесно связаны друг с другом. В Приложении А приведена обобщенная схема взаимосвязи обмена между основными классами соединений: углеводами, жирами и белками.Сказанное не исчерпывает всего многообразия взаимосвязей обмена белков, углеводов и жиров. Между ними существуют более сложные, нежели простое использование в качестве субстратов, формы взаимозависимости.Синтез жиров из углеводов
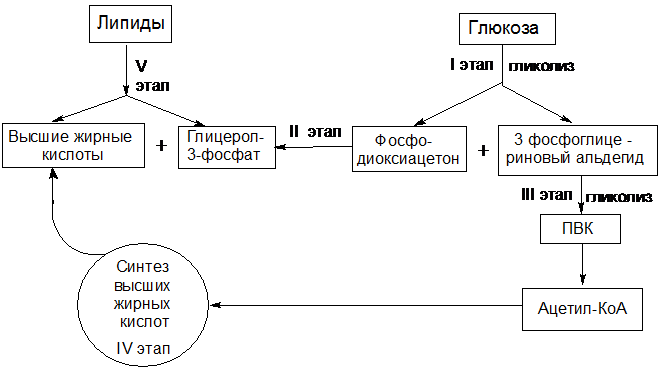
44.
Синтез углеводов из жиров
Процесс синтеза углеводов из жиров можно представить общей схемой:

Рисунок 7 – Общая схема синтеза углеводов из жиров
Один из основных продуктов расщепления липидов – глицерин – легко используется в синтезе углеводов через образование глицеральдегид-3-фосфата и его вступление в глюнеогенез. У растений и микроорганизмов столь же легко используется на синтез углеводов и другой важный продукт расщепления липидов– жирные кислоты (ацетил-КоА), через глиоксилатный цикл.
Но общая схема не отражает всех биохимических процессов, происходящих в результате образования углеводов из жиров.
Поэтому рассмотрим все этапы данного процесса.
Схема синтеза углеводов и жиров более полно представлена на рисунке 8 и происходит в ряд этапов.
1 этап. Гидролитическое расщепление жира под действием фермента липазы на глицерин и высшие жирные кислоты (см. п.1.2). Продукты гидролиза должны, пройдя ряд превращений, превратиться в глюкозу.
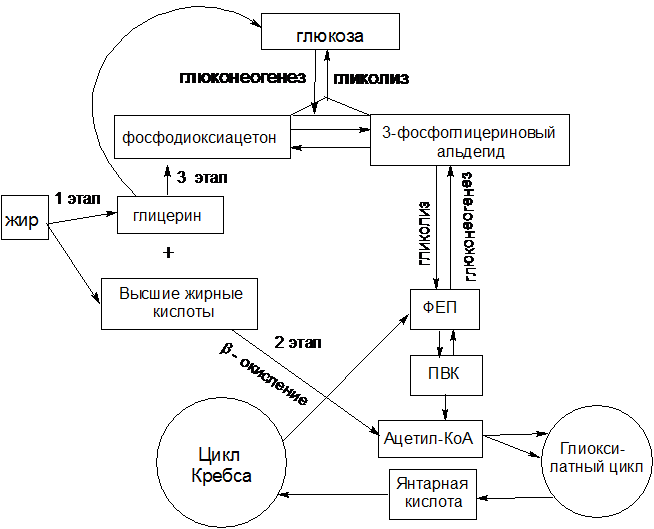
Рисунок 8 – Схема биосинтеза углеводов из жиров
2 этап. Превращение высших жирных кислот в глюкозу. Высшие жирные кислоты, которые образовались в результате гидролиза жира, разрушаются преимущественно путем b-окисления (этот процесс был рассмотрен ранее в разделе 1.2 пункт 1.2.2). Окончательным продуктом этого процесса является ацетил-КоА.
Глиоксилатный цикл
Растения, некоторые бактерии и грибы могут использовать ацетил-КоА не только в цикле Кребса, но и в цикле, получившим название глиоксилатного. Этот цикл играет важную роль в качестве связующего звена в метаболизме жиров и углеводов.
Особенно интенсивно глиоксилатный цикл функционирует в особых клеточных органеллах–глиоксисомах – при прорастании семян масличных растений. При этом происходит превращение жира в углеводы, необходимые для развития проростка семени. Этот процесс функционирует до тех пор, пока у проростка не разовьется способность к фотосинтезу. Когда в конце прорастания запасной жир истощается, глиоксисомы в клетке исчезают.
Глиоксилатный путь специфичен только для растений и бактерий, у животных организмов он отсутствует. Возможность функционирования глиоксилатного цикла связана с тем, что растения и бактерии способны синтезировать такие ферменты, как изоцитратлиаза и малатсинтаза, которые вместе с некоторыми ферментами цикла Кребса участвуют в глиоксилатном цикле.
Схема окисления ацетил-КоА по глиоксилатному пути показана на рисунке 9.
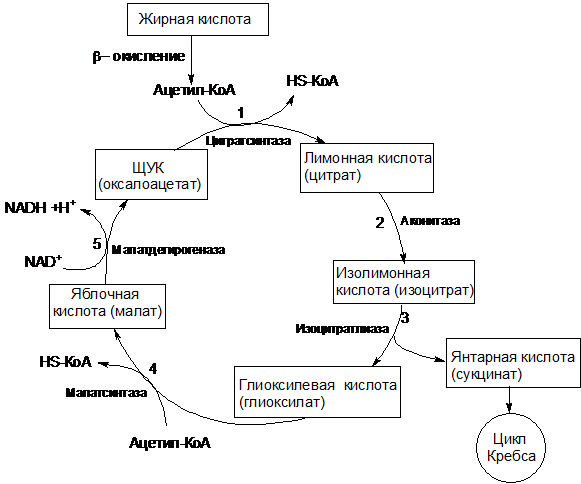
Рисунок 9 – Схема глиоксилатного цикла
Две начальные реакции (1 и 2) глиоксилатного цикла идентичны таковым цикла трикарбоновых кислот. В первой реакции (1) ацетил-КоА конденсируется с оксалоацетатом под действием цитратсинтазы, с образованием цитрата. Во второй реакции цитрат изомеризуется в изоцитрат при участии аконитатгидратазы. Следующие реакции, специфичные для глиоксилатного цикла, катализируются специальными ферментами. В третьей реакции изоцитрат под действием изоцитратлиазы расщепляется на глиоксилевую кислоту и янтарную кислоту:

В ходе четвертой реакции, катализируемой малатсинтазой, глиоксилат конденсируется с ацетил-КоА (второй молекулой ацетил-КоА, вступающей в глиоксилатный цикл) с образованием яблочной кислоты (малат):
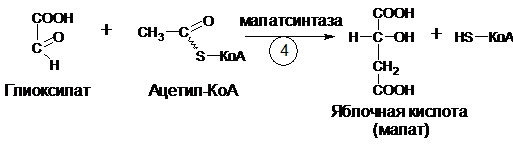
Затем в пятой реакции малат окисляется до оксалоацетата. Эта реакция идентична конечной реакции цикла трикарбоновых кислот; она же является конечной реакцией глиоксилатного цикла, т.к. образовавшийся оксалоацетат вновь конденсируется с новой молекулой ацетил-КоА, начиная тем самым новый оборот цикла.
Образовавшаяся в третьей реакции глиоксилатного цикла янтарная кислота не используется этим циклом, а подвергается дальнейшим превращениям.
Дата добавления: 2018-05-02; просмотров: 619; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!