Ферментативный гидролиз пектина может протекать с участием двух ферментов: пектинэстеразы и полигалактуроназы.

Метоксилированная полигалактуроновая кислота.Пектинэстеразы удаляют метильные группы, гидролизуя сложноэфирные связи находящиеся рядом со свободными карбоксильными группами, т.е. идет реакция:пектин + n Н2О ®n метанол + пектин (менее этерифицированный)Таким образом, пектиновые вещества ответственны за содержание токсичного вещества метанола во фруктовых соках, плодово-ягодных винах.Полигалактуроназа катализирует расщепление α-(1-4)-гликозидной связи, образованной неэтерифицированной галактуроновой кислотой.Препараты, содержащие ферменты, гидролизующие пектиновые вещества, получают обычно из различных плесневых грибов. Эти препараты применяются в пищевой промышленности для осветления фруктовых соков и повышения их выхода, а также для осветления плодовых и виноградных вин, в которых обычно содержится большое количество растворимого пектина, затрудняющего фильтрование и являющегося причиной недостаточной прозрачности вин.
9.Пищевые волокна.Основным источником углеводов в пище человека являются продукты растениеводства. С биохимической точки зрения все углеводы пищи можно подразделить на усвояемые организмом и неусвояемые.Усвояемые углеводы являются основным источником химической энергии в организме, т.е. при их диссимиляции выделяется энергия, необходимая для различных метаболических процессов.
Неусвояемые углеводы не перевариваются в желудочно-кишечном тракте, не всасываются в кишечнике, а если и всасываются, то не вступают в метаболические процессы в организме. К неусвояемым углеводам относятся полисахариды: целлюлоза, гемицеллюлозы, пектиновые вещества; олигосахариды: рафиноза, а также некоторые простые сахара. Пищеварительные секреты слюнной железы, желудка и кишечника не выделяют ферментов, способных расщеплять эти углеводы. Неусвояемые полисахариды образуют группу балластных веществ, называемых пищевыми волокнами. В технологии пищевых продуктов стремятся от них избавиться с целью получения более «рафинированной» пищи. Однако, присутствие их в пищевых продуктах необходимо. Доказано, что недостаток в рационе пищевых волокон является причиной многих желудочно-кишечных заболеваний, атеросклероза и диабета, поскольку они играют существенную роль в поддержании нормальной регуляции питания и метаболизма ряда веществ способствуют лучшему функционированию кишечника. Пищевые волокна, набухая в кишечнике, приобретают способность сорбировать и удалять из организма вредные вещества (токсины), катионы тяжелых металлов, радионуклиды, т.е. выполняют энтеросорбентную функцию. Кроме того, пищевые волокна нормализуют обмен холестерина, способствуют нормальному развитию полезной кишечной микрофлоры.Плоды и овощи - морковь, капуста, свекла – содержат относительно большое количество пищевых волокон – до 1,5%.Не обладая высокой энергетической ценностью, большинство овощей из-за обилия в них клетчатки способствуют раннему и довольно стойкому чувству насыщения. Это свойство пищевых волокон трудно переоценить в комплексе мер профилактики и терапии ожирения.Все это позволило отнести пищевые волокна к одному из резервов здорового долголетия. В суточном рационе взрослого человека должно содержаться 25-30 г пищевых волокон.Наиболее полезны – пектины, лигнин, целлюлоза и гемицеллюлоза пшеничных отрубей, затем идут волокна капусты, затем – морковь.
|
|
|
|
|
|
10.Липиды ( от греч. lipos – жир ) представляют собой группу природных органических соединений, различающихся по своей химической структуре и функциям. Однако, они характеризуются следующими общими признаками: нерастворимостью в воде, растворимостью в органических растворителях (эфире, хлороформе, бензоле), содержанием высших жирных кислот и гидрофобностью. Многие липиды, однако, содержат как минимум одну полярную группу, которая может служить местом связывания с другими компонентами.В организме липиды выполняют пять основных функции:1.Энергетическая. Являются резервными соединениями, основной формой запасания энергии и углерода. При окислении 1г нейтральных жиров (триацилглицеролов) выделяется около 38 кДж энергии.
|
|
|
2. Защитная. Липиды (воски) образуют защитные водоотталкивающие покровы растений, их семян и плодов, и термоизоляционные прослойки у животных организмов.
3. Структурная. Являются главными структурными компонентами клеточных мембран.
4. Поставщик метаболитов–липиды служат предшественниками ряда других биологически активных веществ–витамина Д, стероидных гормонов, желчных кислот, каротиноидов, стеролов и т.д.
5. Регуляторная: 1) производными жирных кислот являются простоглондины–гормоны местного действия. От свойств и структуры мембранных липидов во многом зависит активность мембраносвязанных ферментов; 2) липидами являются жирорастворимые витамины и провитамины (каротины, стеролы), обладая высокой биологической активностью, эти вещества оказывают регулирующее влияние на обмен веществ.
Существует несколько классификаций липидов. Наибольшее распространение получила классификация, основанная на структурных особенностях липидов и их способности к гидролизу.Липиды извлекают из любого растительного материала в виде сложной смеси, и в зависимости от способов и приемов экстрагирования, вида растворителя, различают следующие группы липидов. 1. Свободные липиды, извлекаемые неполярными безводными растворителями (чаще всего используют диэтиловый эфир). При этом в эфирный экстракт переходит не только собственно жир, но и примесь к нему других липидов: свободных высших жирных кислот, высших спиртов, фосфатидов, стеролов, восков, хлорофилла, каротиноидов, жирорастворимых витаминов, т. е. в экстракте находится так называемый сырой жир. В настоящее время принято сырой жир, т.е. все липиды, перешедшие в экстракт диэтилового эфира, называть свободными липидами. Для количественного определения сырого жира используют аппарат Сокслета (см. лаб. практикум). 2. Связанные липиды. Часть липидов может быть связана с белками (липопротеины) и углеводами (гликопротеины). Это вещества извлекаются гидрофильн
|
|
|
ными полярными растворителями или их смесями (хлороформ, этанол, ацетон),
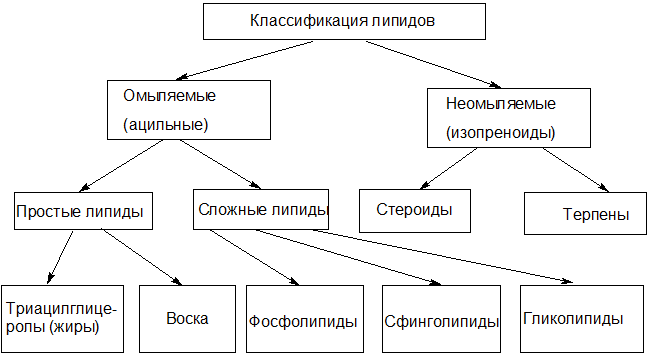
Рисунок 1 – Классификация липидов
которые разрушают непрочные белково-липидные и гликолипидные соединения.
3. Прочносвязанные липиды, извлекаемые после обработки растительного материала спиртовым раствором щелочи при кипячении для разрушения прочных комплексов липидов с нелипидными соединениями; при этом происходит гидролиз и омыление липидов щелочью.
Жирные кислоты
В природе обнаружено свыше 200 жирных кислот, однако более 100 различных жирных кислот идентифицированы в липидах микроорганизмов, растений и животных.Жирные кислоты – алифатические карбоновые кислоты - в организме могут находиться как в свободном состоянии, либо выполнять роль строительных блоков для большинства классов липидов.Все жирные кислоты, входящие в состав жиров, делят на две группы: насыщенные и ненасыщенные, содержащие ненасыщенные жирные кислоты, имеющие две и более двойных связи называют полиненасыщенными.Природные жирные кислоты весьма разнообразны, однако имеют ряд общих черт:- это монокарбоновые кислоты, содержащие линейные углеводородные цепи;- почти все они содержат четное число атомов углерода от 14 до 22, чаще всего встречаются с 16 или 18 атомами углерода; - содержание ненасыщенных жирных кислот в липидах, как правило, выше, чем насыщенных. Двойные связи почти всех природных жирных кислот имеют цис-конфигурацию;- высшие жирные кислоты практически нерастворимы в воде, но их натриевые или калиевые соли, называемые мылами, образуют в воде мицеллы, стабилизируемые за счет гидрофобных взаимодействий. Мыла обладают свойствами поверхностно-активных веществ.
Жирные кислоты отличаются:
- длиной их углеводородного хвоста, степенью их ненасыщенности и положением двойных связей в цепях жирных кислот;- физическими свойствами. Обычно насыщенные жирные кислоты при температуре 22°С имеют твердую консистенцию, тогда как ненасыщенные представляют собой масла. Ненасыщенные жирные кислоты имеют более низкую температуру плавления; - структурной конфигурацией. В насыщенных жирных кислотах углеводородный хвост, в принципе может принимать бесчисленное множество конфигураций вследствие полной свободы вращения вокруг одинарной связи; однако, наиболее вероятной является вытянутая форма, поскольку она энергетически наиболее выгодна. В ненасыщенных кислотах наблюдается иная картина: невозможность вращения вокруг двойной связи (или связей) обусловливает жесткий изгиб углеводородной цепи. В природных жирных кислотах двойная связь, находясь в цис-конфигурации, дает изгиб цепи под углом приблизительно 30°. В жирных кислотах с несколькими двойными связями цис-конфигурация придает углеродной цепи изогнутый и укороченный вид.– химическими свойствами. Полиненасыщенные жирные кислоты быстро окисляются на открытом воздухе. Кислород реагирует с двойными связями с образованием пероксидов и свободных радикалов. Ненасыщенные жирные кислоты участвуют в реакциях присоединения по двойным связям, к ним легко присоединяются галогены.В высших растениях присутствуют в основном пальмитиновая кислота и две ненасыщенные кислоты - олеиновая и линолевая.Стеариновая кислота в растениях почти не встречается, а содержится в значительном количестве (25% и более) в некоторых твердых животных жирах (жир баранов и быков) и маслах тропических растений (кокосовое масло). Лауриновой кислоты много в лавровом листе, миристиновой – в масле мускатного ореха, арахиновой и бегеновой – в арахисовом и соевом маслах. Полиненасыщенные жирные кислоты - линоленовая и линолевая - составляют главную часть льняного, конопляного, подсолнечного, хлопкового и некоторых других растительных масел. Доля ненасыщенных жирных кислот в составе растительных жиров очень высока (до 90%), а из предельных лишь пальмитиновая кислота содержится в них в количестве 10-15%.
В организме человека и животных не могут синтезироваться такие важные кислоты, как линолевая, линоленовая и арахидоновая, и должны поступать в организм с пищей. Эти три кислоты получили название незаменимых жирных кислот. Комплекс этих кислот получил название витамина F. При длительном отсутствии в пище у животных наблюдается отставание в росте, сухость и шелушение кожи, выпадение шерсти. Описаны случаи недостаточности незаменимых жирных кислот и у человека. Так, у детей грудного возраста, получающих искусственное питание с незначительным содержанием жиров, может развиться чешуйчатый дерматит.
Механизм действия витамина F неизвестен, но установлено, что он участвует в регуляции обмена липидов; способствует выведению из организма холестерина, предупреждая и ослабляя атеросклероз; также оказывает благотворное влияние на стенки кровеносных сосудов, повышая их эластичность; являются (в частности арахидоновая кислота) предшественником гормонов - простагландинов, оказывающих влияние на обмен веществ, в частности, ряд простагландинов влияет на деятельность гладких мышц сосудов, в связи с чем их используют для лечения гипертонии, облегчения родов и т. д. Растительные масла не содержат арахидоновой кислоты. Она присутствует в продуктах животного происхождения (яйца, сердце, почки и т. д.) Ежедневно человеку необходимо потреблять в среднем 20-25 г растительного масла и 55-60 г животного.
Масла некоторых растений содержат значительное количество специфических жирных кислот, характерных именно для данных растений. В маслах из семян крестоцветных растений - рапса и горчицы содержится от 42 до 55 % ненасыщенной эруковой кислоты:
СН3-(СН2)7-СН=СН-(СН2)11-СООН
Масло клещевины содержит рицинолевую кислоту - оксикислоту, имеющую гидроксильную группу у 12-го углеродного атома:
СН3-(СН2)5-СН-СН2-СН=СН -(СН2)7-СООН
|
ОН
Триацилглицеролы (жиры)
Нейтральные ацилглицеролы представляют собой сложные эфиры трехатомного спирта глицерина и высших жирных кислот. Простые ацилглицеролы не содержат ионных групп, являются нейтральными липидами, относятся к L-ряду.
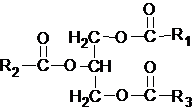
Если жирными кислотами этерифицированы все три гидроксильные группы глицерина, то такое соединение называют триацилглицеролом (триглицерид), если две – диацилглицеролом (диглицерид) и, наконец, если этерифицирована одна группа – моноацилглицеролы( моноглицерид).Триацилглицеролы различаются природой и расположением трех остатков жирных кислот. В зависимости от типа остатков жирных кислот, нейтральные жиры делятся на простые и смешанные. Если во всех трех положениях стоят остатки одной и той же жирной кислоты, то их относят к простым триацилглицеролам, название которых определяется наванием сответствующей жирной кислоты (например, трипальмитин, тристеарин, триолеин и т.д.) если остатки разных жирных кислот, то их относят к смешанным триацилглицеролам (например, 1-олео-2-пальмито-3-стеарин).
Нейтральные ацилглицеролы служат главными составными частями природных жиров и масел, чаще всего это смешанные триацилглицеролы. По происхождению природные жиры делят на животные и растительные. Растительные жиры обычно называют маслами. В зависимости от жирно-кислотного состава жиры и масла по консистенции бывают жидкими и твердыми. Животные жиры (баранье, говяжье, свиное сало, молочный жир) обычно содержит значительное количество насыщенных жирных кислот (пальмитиновой, стеариновой и др.), благодаря чему при комнатной температуре они твердые.Жиры, в состав которых входит много ненасыщенных кислот (олеиновая, линолевая, линоленовая и др.), при обычной температуре жидкие и называются маслами.Жиры, как правило, содержаться в животных тканях, масла - в плодах и семенах растений. Особенно высоко содержание масел (20-60 %) в семенах подсолнечника, хлопчатника, сои, льна. Семена этих культур используются в пищевой промышленности для получения пищевых масел.По способам высыхать на воздухе масла подразделяются на высыхающие (льняное, конопляное), полувысыхающие (подсолнечное, кукурузное), невысыхающие (оливковое, касторовое). Это свойство масел определяется их жирно - кислотным составом. Триацилглицеролы способны вступать во все химические реакции, свойственные сложным эфирам. Наибольшее значение имеет реакция омыления, она может происходить как при ферментативном гидролизе, так и при действии кислот и щелочей. Жидкие растительные масла превращают в твердые жиры при помощи гидрогенизации. Этот процесс широко используется для изготовления маргарина и кулинарного жира.Жиры при сильном и продолжительном взбалтывании с водой образуют эмульсии – дисперсные системы с жидкой дисперсной фазой (жир) и жидкой дисперсионной средой (водой). Однако эти эмульсии нестойки и быстро разделяются на два слоя – жир и воду. Жиры плавают над водой, поскольку их плотность меньше плотности воды (от 0,87 до 0,97).Для получения стойких эмульсий жира в воде необходимо присутствие третьего вещества – эмульгатора, легко адсорбирующегося на поверхности раздела двух фаз. Молекула эмульгатора состоит из двух частей: из углеводородной цепи (гидрофобный хвост) и какой-либо полярной или ионной группы (гидрофильная головка). К таким соединениям относятся мыла, белки, фосфолипиды, соли желчных кислот. При взбалтывании жира с водой в присутствии эмульгатора происходит дробление жира на капли, на которых появляется тончайшая пленка, состоящая из адсорбированных молекул эмульгатора, гидрофобный конец которых погружен в капельку жира, а гидрофильный – в водную фазу.
Таким образом, эмульгатор формирует гидрофильную оболочку вокруг капель жира, образуя мелкодисперсную смесь с водой или эмульсию, которая приобретает свойство стабильности.Эмульгирование имеет большое физиологическое значение при всасывании и усвоении организмом жиров, а также при образовании биологических мембран. Эмульсии широко используются в пищевой промышленности (при приготовлении шоколада, майонеза, маргарина), мыловарении, при изготовлении косметических средств, в производстве красителей.
Воски
Это сложные эфиры высших жирных кислот и высших одноатомных спиртов жирного (реже ароматического ряда).
 Воски являются твердыми соединениями с ярко выраженными гидрофобными свойствами. Природные воски содержат также некоторое количество свободных жирных кислот и высокомолекулярных спиртов. В состав восков входят как обычные, содержащиеся в жирах, – пальмитиновая, стеариновая, олеиновая и др., так и жирные кислоты, характерные для восков, имеющие гораздо большие молекулярные массы, – карноубовая С24Н48О2, церотиновая С27Н54О2,монтановая С29Н58О2 и др.Среди высокомолекулярных спиртов, входящих в состав восков, можно отметить цетиловый – СН3–(СН2)14–СН2ОН, цериловый – СН3–(СН2)24–СН2ОН, мирициловый СН3–(СН2)28–СН2ОН.Воски встречаются как в животных, так и в растительных организмах и выполняют главным образом защитную функцию.В растениях они покрывают тонким слоем листья, стебли и плоды, тем самым, предохраняя их от смачивания водой, высыхания, механических повреждений и поражения микроорганизмами. Нарушение этого налета приводит к быстрой порче плодов при их хранении.Например, значительное количество воска выделяется на поверхности листьев пальмы, произрастающей в Южной Америке. Этот воск, называемый карноубским, является в основном церотиново - мирициловым эфиром:
Воски являются твердыми соединениями с ярко выраженными гидрофобными свойствами. Природные воски содержат также некоторое количество свободных жирных кислот и высокомолекулярных спиртов. В состав восков входят как обычные, содержащиеся в жирах, – пальмитиновая, стеариновая, олеиновая и др., так и жирные кислоты, характерные для восков, имеющие гораздо большие молекулярные массы, – карноубовая С24Н48О2, церотиновая С27Н54О2,монтановая С29Н58О2 и др.Среди высокомолекулярных спиртов, входящих в состав восков, можно отметить цетиловый – СН3–(СН2)14–СН2ОН, цериловый – СН3–(СН2)24–СН2ОН, мирициловый СН3–(СН2)28–СН2ОН.Воски встречаются как в животных, так и в растительных организмах и выполняют главным образом защитную функцию.В растениях они покрывают тонким слоем листья, стебли и плоды, тем самым, предохраняя их от смачивания водой, высыхания, механических повреждений и поражения микроорганизмами. Нарушение этого налета приводит к быстрой порче плодов при их хранении.Например, значительное количество воска выделяется на поверхности листьев пальмы, произрастающей в Южной Америке. Этот воск, называемый карноубским, является в основном церотиново - мирициловым эфиром:
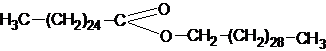
имеет желтый или зеленоватый цвет, очень тверд, плавится при температуре 83-900С, идет на выделку свечей.Среди животных восков наибольшее значение имеет пчелиный воск, под его покровом хранится мед и развиваются личинки пчелы. В пчелином воске преобладает пальмитиново - мирициловый эфир:
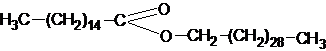
а также высокое содержание высших жирных кислот и различных углеводородов, плавится пчелиный воск при температуре 62-700С.Другими представителями воска животных является ланолин и спермацет. Ланолин предохраняет волосы и кожу от высыхания, очень много его содержится в овечьей шерсти.Спермацет – воск, добывающий из спермацетового масла черепных полостей кашалота, состоит в основном (на 90%) из пальмитиново - цетилового эфира:
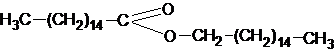
твердое вещество, его температура плавления 41-490С.
Различные воска широко применяют для изготовления свечей, помад, мыла, разных пластырей.
Прогоркание жира
При хранении растительные масла, животные жиры, а также жиросодержащие продукты (масличные жиры, мука, крупа, кондитерские изделия, мясные продукты) под влиянием кислорода воздуха, света, ферментов, влаги приобретают неприятный вкус и запах. Иными словами жир прогоркает.Прогоркание жиров и жиросодержащих продуктов – результат сложных химических и биохимических процессов, протекающих в липидном комплексе.В зависимости от характера основного процесса, протекающего при этом, различают гидролитическоеиокислительное прогоркание, Каждый из них может быть разделен на автокаталитическое (неферментативное) и ферментативное (биохимическое) прогоркание.При гидролитическомпрогоркании происходит гидролиз жира с образованием глицерина и свободных жирных кислот.
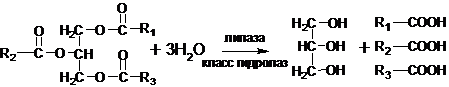
Неферментативный гидролиз протекает с участием растворенной в жире воды и скорость гидролиза жира при обычных температурах невелика. Ферментативный гидролиз происходит при участии фермента липазы на поверхности соприкосновения жира и воды и возрастает при эмульгировании.В результате гидролитического прогоркания увеличивается кислотность, появляется неприятный вкус и запах, особенно при гидролизе жиров (молочного жира, кокосового и пальмового масел), содержащих низко- и среднемолекулярные кислоты, такие как масляную, валериановую, капроновую). Высокомолекулярные кислоты вкуса и запаха не имеют и повышение их содержания не приводит к уменьшению вкуса масел.В семенах растений содержится фермент триацилглицерол–липаза, она особенно активна в семенах клещевины и сои. Участие этого фермента в гидролизе жира зерна и продуктов его переработки (мука, крупа, макароны) приводит к снижению их потребительских свойств и способности к длительному хранению. Наиболее распространенным видом порчи жиров в процессе хранения является окислительное прогоркание. В первую очередь окислению подвергаются свободные, а не связанные в триацилглицеролах ненасыщенные жирные кислоты. Процесс окисления может происходить неферментативным и ферментативным путями.В результате неферментативного окисления кислород присоединяется к ненасыщенным жирным кислотам по месту двойной связи с образованием циклической перекиси, которая распадается с образованием альдегидов, придающих жиру неприятный запах и вкус:
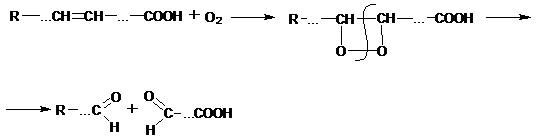
Так же в основе неферментативного окислительного прогоркания лежат цепные радикальные процессы, в которых участвуют кислород и ненасыщенные жирные кислоты.

Под влиянием перекисей и гидроперекисей происходит дальнейший распад жирных кислот, также образуются альдегиды, кетоны и другие неприятные на вкус и запах вещества вследствие чего жир прогоркает. При ферментативном окислении этот процесс катализируется ферментом липоксигеназой с образованием перекисей и гидроперекисей.Специфичность этого фермента состоит в том, что действию фермента подвергаются лишь те полиненасыщенные жирные кислоты, которые содержат цис-цис-1,4-пентадиеновую группу (линолевая, линоленовая, арахидоновая). Процесс начинается с отщепления атома водорода у 3 атома углерода пентадиеновой группы жирной кислоты. Образовавшийся свободный радикал перемещается к 5-му атому углерода с одновременным перемещением двойной связи в сопряженное положение, которая при этом из цис-цис формы переходит в цис-транс изомер, в результате данного процесса идет образование гидроперекиси:

Жирные кислоты с цис-транс или транс-транс-конфигураций двойных связей ферментом не окисляется.
Липоксигеназа широко распространена в растительном жире. Она найдена в пшенице и других злаках, в семенах масличных и бобовых растений, особенно велико ее содержание в соевой муке. В результате действия этого фермента происходит прогоркание пищевых продуктов (муки, крупы, макарон и др.), образующиеся под воздействием гидроперекиси жирных кислот могут далее окислять ненасыщенные жирные кислоты, каротин, витамин А, аминокислоты и аскорбиновую кислоту. Таким образом снижается ценность продукта и изменяется цвет. В этом проявляется ее отрицательное действие.Активность липоксигеназы необходимо учитывать в некоторых технологиях, т.к. она может влиять на потребительские свойства готового продукта.Например, при производстве макарон требуется специальная макаронная мука, вырабатываемая из твердых сортов пшеницы, с низкой активностью липоксигеназы в муке, получается бледные макароны, из-за окисления ею пигментов муки (каротиноидов, флавоноидов и т.д.). Потребительские свойства (цвет) таких макарон не высокие.Поэтому при переработке сырья и выработке из него продуктов важны ингибиторы липоксигеназы, которые взаимодействуют с образующимися свободными радикалами. Эти ингибиторы прекращают процесс окисления, обрывая цепь превращений радикалов. Таким ингибитором является используемый в качестве антиоксиданта токоферол – витамин Е.Липоксигеназа может выполнять и положительную роль. При слабом ее действии небольшие количества гидроперекисей жирных кислот (окисляя группы –SH в белках и образуя дисульфидные группы –S–S–) «укрепляют» клейковину, ускоряют процесс «созревания» пшеничной муки, улучшает ее хлебопекарные достоинства.Разработана технология выработки хлеба из слабой пшеничной муки с использованием липоксигеназы. К пшеничной муке добавляют соевую или гороховую муку (источник липоксигеназы) и растительное масло (источник ненасыщенных жирных кислот). В результате разностороннего действия фермента (укрепление клейковины и обесцвечивание пигментов муки) улучшается цвет мякиша (он становится светлее) и увеличивается объем.Под действием ферментов липазы и липоксигеназы изменяется качество жиров и масел, которое характеризуется следующими показателями и числами:Кислотное число (К.ч.) – это количество миллиграммов гидроксида калия, необходимого для нейтрализации свободных жирных кислот в 1 г жира.При хранении масла наблюдается гидролиз триацилглицеролов, это приводит к накоплению свободных жирных кислот, т.е. возрастанию кислотности. Повышение К.ч. указывает на снижение его качества. Кислотное число является гостированным показателем масла или жира.Йодное число – это количество граммов йода, присоединившемуся по месту двойных связей к 100 г жира:
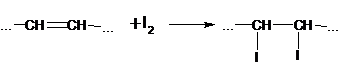
Йодное число позволяет судить о степени насыщенности масла (жира), о склонности его к высыханию, прогорканию и другим изменениям, происходящим при хранении. Чем больше содержится в жире ненасыщенных жирных кислот, тем выше йодное число. Уменьшение йодного числа в процессе хранения масла является показателем его порчи.
Перекисное число (П.ч.) показывает количество перекисей в жире, выражают его в процентах йода, выделенного из йодистого калия перекисями, образовавшимися в 1 г жира.

В свежем жире перекиси отсутствуют, но при доступе воздуха они появляются сравнительно быстро. В процессе хранения перекисное число увеличивается.
Число омыления (Ч.о.) – это количество миллиграммов гидроксида калия , необходимое для нейтрализации свободных и омыления связанных с глицерином жирных кислот в 1 г жира.
Фосфолипиды
Фосфолипиды являются структурными компонентами клеточных мембран и в небольшом количестве обнаружены в составе запасных отложений. Они легко образуют комплексы с белками и в виде фосфолипопротеинов присутствуют во всех клетках живых существ.Фосфолипиды найдены в животных и растительных организмах. Особенно много их содержится в нервных тканях, в сердце, печени животных. Много фосфолипидов в семенах растений, особенно в семенах масличных и бобовых культур.Фосфолипиды – твердые вещества жироподобного вида; они бесцветны, но быстро темнеют на воздухе вследствие окисления ненасыщенных жирных кислот. Они хорошо растворимы в большинстве неполярных органических растворителей (бензол, хлороформ, петролейный эфир) и масле. В воде они не растворимы, но могут образовывать эмульсии или коллоидные растворы.Фосфолипиды – сложные эфиры многоатомных спиртов с высшими жирными кислотами, содержащие в качестве добавочных групп остатки фосфорной кислоты и азотистых оснований.Из многоатомных спиртов в составе различных фосфолипидов найдены глицерин, миоинозит и сфингозин.В соответствии с этим фосфолипиды делят на три группы: глицерофосфолипиды, ионозитфосфолипиды и сфингофосфолипиды. В качестве высших жирных кислот в молекулах фосфолипидов содержится пальмитиновая, стеариновая, линолевая, линоленовая и арахидоновая кислоты.Наиболее распространенными в природе являются глицерофосфолипиды и сфингофосфолипиды.
Глицерофосфолипиды
Глицерофосфолипиды являются производными фосфатидной кислоты. В их состав входят глицерин, жирные кислоты, фосфорная кислота и обычно азотсодержащее соединение. Общая формула глицерофосфолипидов выглядит так:
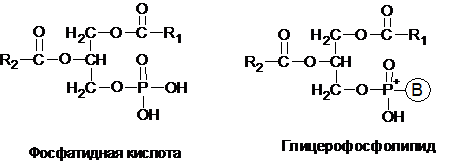
В этих формулах R1 и R2 – радикалы высших жирных кислот (как правило, R2–это ненасыщенная жирная кислота), а В – радикал азотистого основания.
В зависимости от характера азотистого основания среди фосфотидов различают фосфотидилхолины (лецитины), если фосфорная кислота соединена эфирной связью с азотистым основанием – холином [НО–СН2–СН2–N+(CН3)3]; фосфатидилэтаноламины (кефалины), если азотистым основанием служит этаноламин (HO-CH2-CH2-H3N+); фосфатидилсерины, если азотистым основанием является аминокислота серин (HO-CH2-CH-H3N+):
|
СОО-
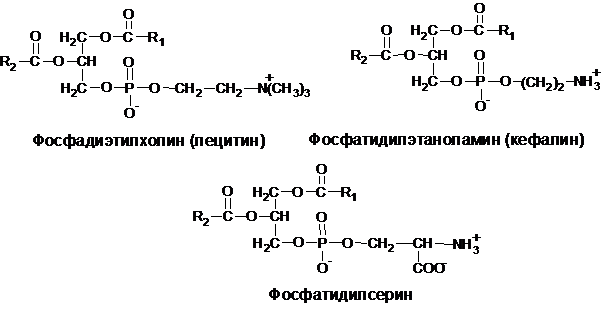
Три первых вида азотосодержащих фосфатидов могут переходить друг в друга, т. к. они отличаются лишь строением азотистых оснований, между которыми возможна, например, такая генетическая связь:

В состав некоторых глицерофосфолипидов вместо азотосодеращих соединений входит не содержащий азота шестиуглеродный циклический спирт инозит, названный также инозитолом. Эти липиды называются инозитфосфолипиды или фосфатидилинозитолами:
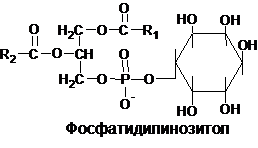
Фосфотидилинозитолы обнаружены у животных, растений и микроорганизмах. В животном организме найдены в мозге, печени и легких.
Сфингофосфолипиды
Особую группу составляют фосфолипиды, у которых вместо глицерина содержится аминоспирт сфингозин, такие фосфолипиды называются сфингофосфолипидами.К наиболее распространенным сфинголипидам относят сфингомиелин.
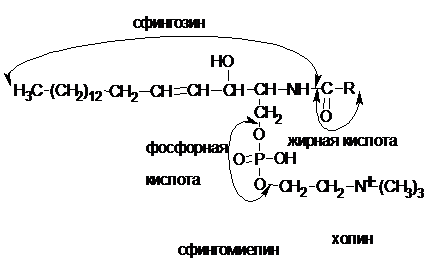
В основном они находятся в мембранах животных и растительных клеток. Особенно богата ими нервная ткань, также сфингомиелины обнаружены в ткани почек, печени и других тканей.Молекулы фосфолипидов, особенно глицерофосфолипидов и сфингофосфолипидов, обладают выраженной полярностью. В структуре фосфолипидов можно выделить два фрагмента: полярную голову, образованную фосфорной кислотой и азотосодержащим соединением (холин, этаноламин, серин) и гидрофобные «хвосты», образованные остатками высших жирных кислот.Благодаря такому строению фосфолипиды обладают следующими свойствами:- в растворах образуют слоистые структуры, которые играют очень важную роль в построении клеточных мембран. Фосфолипиды, образуя структурный матрикс мембран в виде двойных липидных слоев, являются основой любой биологической мембраны;- образуя комплексы с белками клеточных мембран, регулируют процессы транспорта ионов и других органических веществ через мембрану, обеспечивая нормальный ход обменных процессов в клетке;- являются природными антиоксидантами и предохраняют масла от окисления, легко окисляясь сами;
- хорошие поверхностно-активные вещества (ПАВ), способные оказывать влияние на структурно-механические свойства клейковины, улучшая хлебопекарные достоинства пшеничной муки;- прекрасные эмульгаторы (особенно лецитин) и широко используются в пищевой промышленности при изготовлении шоколада, маргарина, майонеза.
Гликолипиды
Гликолипиды в отличии от фосфолипидов не содержат фосфорной кислоты, а вместо азотистого основания в их состав входят углеводы, чаще всего галактоза или ее производные. Из спиртов гликолипиды содержат глицерол или сфингозин, а также остатки жирных кислот.
Среди гликолипидов особенно широко распространены галактозилацилглицеролы.
Эти соединения содержатся в самых различных растительных тканях. Они обнаружены в митохондриях, хлоропластах и локализованы в мембранах; содержатся в водорослях, некоторых фотосинтезирующих бактериях.
Главной формой гликолипидов в животных тканях, особенно в нервной ткани, в частности в мозге, являются гликосфинголипиды. Последний содержит церамид, состоящий из спирта сфингозина и остатка жирной кислоты, и один или несколько остатков сахаров. Важнейшими гликосфинголипидами являются цероброзиды и ганглиозиды.
Простейшими цероброзидами являются галактозилцерамиды и глюкозилцерамиды. В состав галактозилцерамидов входит Д-галактоза, которая связана эфирной связью с гидроксильной группой аминоспирта сфингозина. Кроме того, в составе галактозилцерамида имеется жирная кислота. Чаще всего лигноцериновая, нервоновая или цереброновая кислота, т.е. жирные кислоты, имеющие 24 углеродных атома.

Рисунок 5 – Структура галактозилцерамида
Существуют сульфогалактозилцерамиды, которые отличаются от галактозилцерамидов наличием остатка серной кислоты, присоединенного к третьему углеродному атому гексозы.
Глюкозилцерамиды в отличие от галактозилцерамидов вместо остатка галактозы имеется остаток глюкозы.
Более сложными гликосфинголипидами являются ганглиозиды. Одним из простейших ганглиозидов является гематозид, выделенный из стромы эритроцитов. Он содержит церамид, по одной молекуле галактозы, глюкозы и N-ацетилнейраминовой кислоты. Ганглиозиды в большом количестве находятся в нервной ткани. Они выполняют рецепторные и другие функции.
Стероиды
Стероиды – широко распространенные в природе соединения. Это производные тетрациклических тритерпенов. Основу их структуры составляет циклопентанпергидрофенантреновое ядро:

К стероидам относят стерины (стеролы) – высокомолекулярные циклические спирты и стериды – сложные эфиры стеринов и высших жирных кислот. Стериды не растворяются в воде, но хорошо растворимы во всех жировых растворителях и входят в состав сырого жира. Стериды образуют омыляемую фракцию липидов. Стерины же при омылении жира остаются в неомыляемой фракции, составляя наибольшую ее часть.В организме человека и животных главным представителем стеринов (стеролов) является холестерин:
 Холестерин играет важную роль в жизнедеятельности животного организма:
Холестерин играет важную роль в жизнедеятельности животного организма:
– участвует в построении биологических мембран. Находясь в составе мембран клеток, вместе с фосфолипидами и белками обеспечивает избирательную проницаемость клеточной мембраны, оказывает регулирующее влияние на состояние мембраны и на активность связанных с ней ферментов;– является предшественником образования в организме желчных кислот, а также стероидных гормонов. К этим гормонам относятся тестостерон (мужской половой гормон), эстрадиол (один из женских гормонов), альдестерон (образующийся в коре надпочечников и регулирующий водно – солевой баланс);– является провитамином витаминов группы Д. Холестерин под действием УФ-лучей в коже превращается в витамин Д3 (холекальциферол), который в свою очередь служит предшественником гормона, участвующего в регуляции обмена кальция и минерализации костной ткани. Так же нужно отметить, что при нарушении обмена веществ холестерин откладывается на стенках кровеносных сосудов, приводя к тяжелой болезни – атеросклерозу.
В растениях и дрожжах содержится эргостерин (эргостерол):
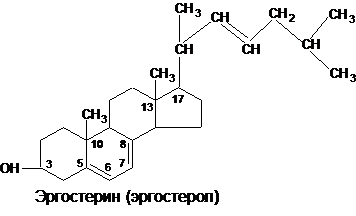
При облучении эргостерола УФ из него образуется витамин Д2 (эргокальциферол). Для промышленного изготовления витаминов группы Д (антирахитические витамины) используют дрожжи, они содержат свыше 2% стеридов и стеролов на сухое вещество.В растительных маслах (соевое. кукурузное, масла пшеничных зародышей) содержится обычно от двух до четырех различных стеролов, отличающихся друг от друга количеством, расположением двойных связей и строением боковой цепи, причем обязательной составной частью является β-ситостерол:
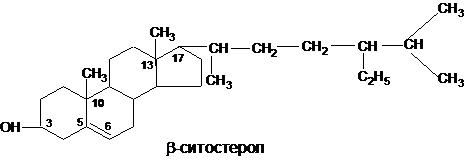
В кукурузе доля β-ситостерола составляет 86% от всех стеролов, а в пшенице – 66%.
Терпены
В основе строения терпенов находится молекула изопрена:
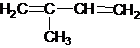
Это мономер, из которого построены олигомерные или полимерные цепочки неомыляемых липидов. Терпены, молекулы которых представляют собой соединения из 2, 3, 4, 6, 8 молекул изопрена, называют соответственно моно-, сескви-, ди-, три- и тетратерпенами. Молекулы терпенов могут иметь линейную или циклическую структуру, содержать гидроксильные, карбонильные и карбоксильные группы.Монотерпены.это летучие жидкие вещества с приятным запахом. Они являются основными компонентами душистых эфирных масел, получаемых из растительных тканей – цветов, листьев, плодов.В качестве типичного представителя алифатических монотерпенов является мирцен. От 30 до 50% мирцена содержится в эфирном масле хмеля. Представителями кислородных производных алифатических терпенов являются линалоол, гераниол и цитронеллол. Все они представляют спирты. Линалоол содержится в цветках ландыша, в апельсиновом и кориандровом масле. По-видимому, аромат персиков обусловлен различными сложными эфирами линалоола – уксуснокислым, муравьинокислым и др. Гераниол встречается в масле эвкалипта. Цитронеллол обладает запахом розы и содержится в розовом, гераниевом и других маслах.Среди моноциклических терпенов наиболее распространенным и важным являются лимонен, ментол, карвон. Лимонен содержится в скипидаре, тминном масле; ментол составляет главную (до 70%) эфирного масла перечной мяты, а карвон содержится в эфирных маслах тмина и укропа.Сесквитерпены.Эта группа терпенов также входит в составэфирных масел. Одно из наиболее интересных соединений – ароматический сесквитерпен димер госсипол – специфический пигмент хлопковых семян.
Дитерпены. Наиболее широко представлены соединениями, входящими в состав многих биологически важных соединений. Так, дитерпеновый спирт фитол входит в состав хлорофилла.
Хлорофилл – это пигмент, придающий растениям зеленый цвет. Он содержится в листьях и стеблях, в колосьях и зернах. Хлорофилл находится в особых образованиях протоплазме, называемых хлоропластами. В растениях существуют два вида хлорофилла: хлорофилл а (сине-зеленый) и хлорофилл в (желто-зеленый)

Большой интерес представляет сходство строения хлорофилла с красящим веществом крови гемином. В состав хлорофилла и гемина входят четыре остатка пиррола, соединенных в виде порфиринового ряда, которое в гемине связано с железом, а в хлорофилле – с магнием. Хлорофилл принимает активное участие в процессе фотосинтеза. В результате этого процесса диоксид углерода под влиянием солнечного света поглощенного хлорофиллом, восстанавливается до гексозы и выделяется свободный кислород. Фотосинтез – это единственный процесс, в ходе которого лучистая энергия солнца в виде химических связей запасается в органических соединениях.
Дитерпеновые цепи входят в состав витаминов Е и К1; витамин А – это моноциклический дитерпен. Трициклическим дитерпеном служит абиетиновая кислота – главный компонент смоляных кислот, известный в технике как канифоль. Натриевые соли канифоли – это один из компонентов хозяйственногомыла.Многие дитерпены являются компонентами эфирных масел – камфорен, каурен, стевиол и агатовая кислота.
Тритерпены. Представлены наиболее известным тритерпеном скваленом. Сквален – исходное соединение, из которого у животных и дрожжей, синтезируются стероиды, например, холестерол. Тритерпеновая цепь входит в состав витамина К2. К более сложным тритерпенам относятся лимонин и кукурбитацин А – соединения, обуславливающие горький вкус лимона и тыквы.
Тетратерпены. Это пигменты – каротиноиды. Они придают растениям желтую или оранжевую окраску разных оттенков. Наиболее известные представители каротиноидов – каротин, лютеин, цеаксантин и криптоксантин.Каротины впервые выделены из моркови (от лат. «карота» – морковь). Известно три типа каротинов: α-, β- и γ-каротины, отличающиеся как по химическому строению, так и по биологическим функциям. Наибольшей биологической активностью обладает β-каротин, так как он содержит два β-иононовых кольца и при его гидролитическом распаде под действием фермента каротиназы образуется две молекулы витамина А1:
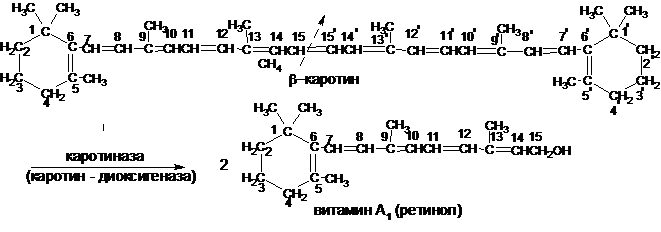
При гидролитическом расщеплении α- и γ-каротина образуется по одной молекуле витамина А, так как они содержат по одному β-иононовому кольцу. Степень усваяемости каротиноидов и свободного витамина А зависит от содержания жиров в пище. β-Каротин придает моркови, тыкве, апельсинам, персикам и другим овощам и фруктам характерный для них цвет. Каротины наряду с хлорофиллом содержатся во всех зеленых частях растений.Лютеин – желтый пигмент, содержащийся наряду с каротинами в зеленых частях растений. Окраска семян желтой кукурузы зависит от присутствующих в них каротинов и каротиноидов, получивших название цеаксантина и криптоксантина. Окраска плодов томата обусловлена каротиноидом ликопином.Лютеин, цеаксантин и криптоксантин также обнаруживают активность витамина А.Каротиноиды играют большую роль в обмене веществ у растений, участвуя в процессе фотосинтеза. Также каротиноиды имеют большое значение в пищевой промышленности. Пигментация каротиноидами зерна хлебных злаков влияет на их технологическое достоинство. Кремовый цвет, характерный для хорошей пшеничной муки, объясняется главным образом содержанием каротиноидов. Пигментирование зерна особенно высоко ценится у твердой пшеницы, идущей на выработку макаронной муки и манной крупы. Каротиноиды используются как природные красители в кондитерской и масло – жировой промышленностях.
Пентатерпены. Представители этой группы – убихинон Q, пластохиноны выполняют роль переносчиков электронов в окислительно – восстановительных реакциях, связанных с синтезом АТФ.
Биологические мембраны
Мембраны – это универсальная форма структурной организации живой материи. Биологические мембраны или биомембраны, формируют внешнюю границу клетки, делят объем цитоплазмы клетки на отсеки, или компарменты, в которых осуществляются метаболические реакции.
Мембрана содержит липиды и белки, а также небольшое количество углеводов в составе гликосфинголипидов и гликопротеинов. Биомембраны имеют жидкостно-мозаичную структуру, согласно которой мембрана состоит из бислоя сложных липидов (фосфолипидов, гликолипидов, стеролов), в которых находятся глобулярные белки.
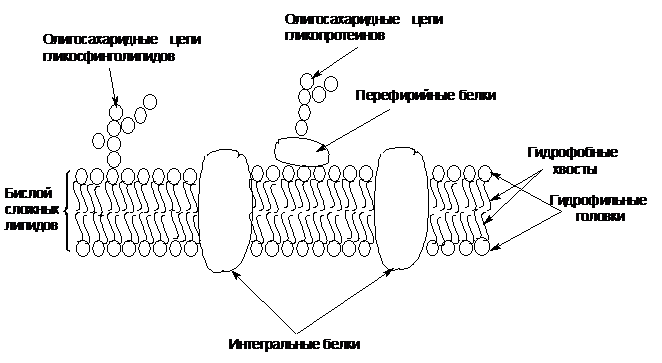 Рисунок 6 – Схема строения двойного липидного слоя в биомембране
Рисунок 6 – Схема строения двойного липидного слоя в биомембране
Липиды в бислое расположены так, что их гидрофобные хвосты (углеводородные остатки жирных кислот) направлены внутрь мембраны на встречу друг другу и частично перекрываются, обе поверхности бислоя составлены гидрофильными головками липидов – фосфолипидов. Белки, встроенные в липидный бислой мембраны, могут быть перефирийными или интегральными. Перефирийные белки связаны с полярной поверхностью бислоя относительно слабыми водородными связями и легко извлекаются из мембраны. Интегральные белки погружены в липидный слой, иногда пронизывают ее насквозь. Удерживаются они значительно прочнее за счет большего числа гидрофобных взаимодействий, водородных и других связей.
Биомембраны ассиметричны, их наружная поверхность отличается по свойствам от внутренней поверхности. Ассиметричность биомембраны обусловлена главным образом различием в природе белков, располагающихся на внешней и внутренней поверхности биомембраны.
Все биомембраны имеют принципиально одинаковую структуру, отличаясь только в деталях. Они могут иметь различные по химическому составу липиды в бислое, разные жирно-кислотные остатки в составе фосфолипидов, разные типы (по химическому строению, гидрофильным и гидрофобным свойствам) встроенных белков. Соотношение липид:белок в биомембране также может варьировать.
Биомембраны представляют собой не просто оболочки клеток и субклеточных структур, но и выполняют ряд важнейших разнообразных функций:
1. Играют роль механического барьера. Участвуют в формировании клеточных структур, ограничивают их от внешней среды;
2. Выполняют транспортную функцию. Транспортные системы, локализованные в мембранах, обеспечивают перенос определенных соединений. В качестве транспортных систем в мембранах выступают специфические транспортеры. К ним относят ионофоры, которые увеличивают скорость диффузии ионов через мембрану, и представляют собой токсины, синтезирующиеся грибами и бактериями. Также специфическими транспортерами являютсябелки. Перенос небольших молекул и ионов через мембраны обеспечивается следующими типами интегральных мембранных белков: каналами и порами; пассивными и активными транспортерами. Белки, формирующие каналы и поры, содержат гидрофильный проход, который позволяет молекулам и ионам с подходящим размером, зарядом и геометрической конфигурацией проникать через мембрану в каком-либо направлении. Активные и пассивные транспортеры, в отличие от каналов и пор, специфически связывают субстрат и транспортируют его через мембрану, изменяя свою конформацию. Пассивные транспортеры способны перемещать молекулы по концентрационному градиенту и не требуют затрат энергии. Для функционирования активных транспортеров, переносящих субстраты против концентрационного градиента, необходимы затраты энергии. При первичном активном транспорте энергия поставляется непосредственно за счет гидролиза АТФ, энергии света или электронного транспорта. Вторичный активный транспорт обеспечивается с помощью ионных градиентов.
3. Выполняют разделительную функцию. Биомембраны регулируют поток веществ внутрь клетки и из нее, что важно для сохранения постоянства внутриклеточной среды.
4. Имеют избирательную проницаемость,которая основана на системах специфичности транспортных белков, способных узнавать определенное соединение и проводить его через мембрану.
5. Важнейшая функция биомембран – генерирование биоэлектрического потенциала, аккумулирование и трансформация энергии. Мембраны являются теми клеточными структурами, благодаря которым возможна трансформация осмотической и электрической форм энергии в энергию макроэргической фосфатной связи (~) АТФ. Так, процессы фотосинтеза и дыхания осуществляются, соответственно, в мембранах хлоропластов и митохондрий. Например, при дыхании энергия освобождается в мембранах митохондрий и используется в процессах биосинтеза различных веществ.
6. Выполняют рецепторную функцию. В связи с наличием в мембранах специфических белковых рецепторов клетки способны воспринимать внешние сигналы и в ответ на них перестраивать свой обмен веществ.
7. Выполняют адгезивную функцию, благодаря которой клетки способны взаимодействовать друг с другом, что существенно, в частности, при формировании тканей.
8. Обеспечивают важнейшие метаболические процессы. Это связано с тем, что в большинстве мембран содержаться ферменты и ферментные комплексы, которые способны осуществлять различные метаболические процессы.
21.Биологическое окисление наз.совокупность овр происходящих в биологических объектах и обеспечивающих их энергией и метаболитами для осущ. процесса жизнедеятельности.Существуют два типа окисления свободное окисление,несопряженное фосфорилирование с адф и и ситезом атф.При свободном окислении освобождающиеся при разрыве хим связи энергия переходит в тепловую энергию и рассеивается.по типу свободного окисления идут все окислительно-восст. реакции с участием ферментов относящиееся к классу оксидоредуктаз.виды фосфорилирования:1)фотосинтетическое. в этом процессе атф синтезируется за счет солнечной энергии,кот. исп.при фотосинтезе.2)субстратное фосфорилирование.если фосфорилирование адф происходит засчет разрыва макроэргической связи в субстрате S+E=ES=ES0=P+E
Окисл. фосфор.если атомы водорода в коферментах дегидрогеназ принимающих участие в окислении Sсубстратов передаются в митохондриальную дыхательную цепь,где происходит перенос электронов и пртонов на кислород молекулярный.и связано всё это с фосфорилированием адф и синтезом атф
в результате окисления одной молекулы НАДН+ в мдц выделяется три молекулы атф.в результате окисление одной молекулы ФАДН2 В МДЦ выделяется две молекулы атф.
22.Обмен веществ и энергии составляет сущность жизнедеятельности любого организма. Обмен веществ представляет собой непрерывный, самосовершающийся и саморегулируемый круговорот веществ, протекающий в процессе существования живой материи и сопровождающийся ее постоянным самообновлением. Иными словами обмен веществ есть диалектическое единство противоположных процессов: ассимиляции (синтеза) и диссимиляции (распада), т.е. единство противоположных процессов питания и выделения, усвоения и разрушения, синтеза и распада.
Обмен веществ невозможен без сопутствующего ему обмена энергии. Каждое органическое соединение, входящее в состав живой материи, обладает определенным запасом потенциальной энергии, за счет которой может быть совершена работа. Эту энергию называют свободной энергией. Уровни свободной энергии индивидуальных исходных веществ и продуктов реакции, как правило, различны, вследствие чего в процессе преобразования веществ происходит перераспределение свободной энергии между компонентами реакционной смеси, т.е. протекает обмен энергией между веществами. Главнымиматериальными носителями свободной энергии в органических веществах являются химические связи между атомами, поэтому при преобразовании химических связей в молекуле уровень свободной энергии соединения изменяется. Нормальными считаются связи, при возникновении или распаде которой уровень свободной энергии составляет около 12,5 кДж/моль. Однако, при новообразовании и распаде некоторых связей уровень свободной энергии в молекулах ряда органических веществ изменяется в гораздо большей степени и составляет 25-50 кДж/моль и более. Такие соединения называются макроэргическими, а связи, при образовании которых наступают столь крупные изменения в энергетическом балансе вещества – макроэргическимисвязями. В отличие от обычных они обозначаются значком (~). В качестве примеров можно привести АТФ, глицерол-3-фосфат, коэнзим А, креатин-фосфат и др.
Макроэргические вещества выполняют функцию и доноров, и акцепторов энергии в обмене веществ, они служат как аккумуляторами, так и проводниками энергии в биохимических процессах. Кроме того, им свойственна роль трансформаторов энергии, так как они способны преобразовывать стационарную форму энергии химической связи в мобильную, т.е. в энергию возбужденного состояния молекулы. Наиболее важным макроэргическимсоединением в организме является аденозинтрифосфорная кислота – АТФ:
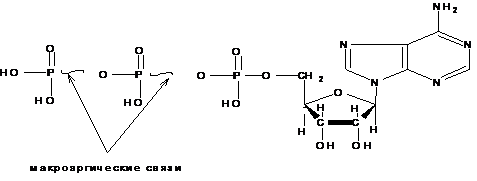
Брожение и дыхание
Окисление органических соединений, как уже указывалось, является основным источником энергии в живом организме в целом, и в клетке, в частности.
Окисление органических соединений (углеводов, жиров, белков) в организме может протекать двумя путями: в присутствии кислорода воздуха – этот тип окисления называется аэробным, или дыханием, и в отсутствии кислорода – этот тип окисления называется анаэробным, или брожением.
Общее уравнение дыхания глюкозы, например:
С6Н12О6 + 6О2® 6СО2 + 6Н2О + 2780 кДж.
Различают три основных типа брожения:
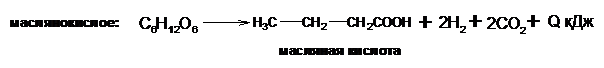
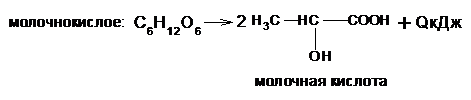
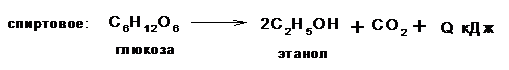
Кроме этих основных видов брожения в природе существуют пропионовокислое, уксуснокислое, ацетоновое и другие виды, но они являются определенным сочетанием основных типов брожение и находят применение или сопровождают технологические процессы в различных отраслях промышленности.
Здесь мы приводим суммарные уравнения реакций аэробного и анаэробного окисления глюкозы. Но процесс окисления, в целом, представляет собой множество последовательно протекающих ферментативных реакций в клетках.
Конечной целью обоих этих процессов является:
а) синтез АТФ за счет утилизации энергии, аккумулированной в углеводах;
б) получение пластических соединений, являющихся исходным материалом для синтеза компонентов клетки.
Гликолиз
Гликолиз–универсальный и основной процесс катаболизма углеводов для большинства организмов, это последовательность реакций, приводящих к превращению глюкозы в пируват с одновременным образованием АТФ.
Важнейшими моносахаридами, катаболизм которых осуществляется по гликолитическому пути, являются Д-глюкоза и Д-фруктоза. Однако и другие моносахариды способны распадаться по пути гликолиза, поскольку они легко превращаются в эти сахара. Гликолиз–процесс анаэробный, однако он может протекать как в отсутствии, так и в присутствии кислорода. Он является ключевым метаболическим путем, генерирующим энергию в форме АТФ в клетках, где отсутствует фотосинтез.
Исследования химизма гликолиза показали, что начальные этапы процессов брожения и дыхания имеют общий путь. Это открытие было уникальным, потому что оно вскрывало существование внутреннего единства живой материи. При дыхании у аэробных организмов гликолиз предшествует циклу трикарбоновых кислот и цепи переноса электронов. Пируват проникает в митохондрии, где он полностью окисляется до СО2, в результате чего с высокой эффективностью из гексозы извлекается свободная энергия. При брожении, в анаэробных условиях (дрожжи, молочнокислые бактерии), пируват превращается в продукты брожения (этанол, лактат).
Все реакции гликолиза протекают в цитоплазме клетки и катализируются десятью различными ферментами.
Первой ферментативной реакцией гликолиза является фосфорилирование, т.е. перенос остатка ортофосфата на глюкозу за счет АТФ. Реакция катализируется ферментом гексокиназой, относящимся к классу трансферазы:

 В дальнейшем остаток фосфорной кислоты
В дальнейшем остаток фосфорной кислоты 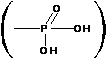 будем обозначать символом: .
будем обозначать символом: .
Второй реакцией гликолиза является превращение глюкозо-6-фосфата под действием фермента глюкозо-6-фосфат-изомеразы во фруктозо-6-фосфат:
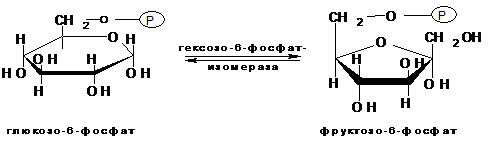
Третья реакция катализируется ферментом фосфофруктокиназой (класс трансферазы):
 Четвертую реакцию гликолиза катализирует фермент альдолаза (класс лиазы). Под влиянием этого фермента фруктозо-1,6-бисфосфат расщепляется на две фосфотриозы:
Четвертую реакцию гликолиза катализирует фермент альдолаза (класс лиазы). Под влиянием этого фермента фруктозо-1,6-бисфосфат расщепляется на две фосфотриозы:
 Пятая реакция – это реакция изомеризации триозофосфатов, катализируется ферментом триозофосфатизомеразой:
Пятая реакция – это реакция изомеризации триозофосфатов, катализируется ферментом триозофосфатизомеразой:

В последующие реакции гликолиза может непосредственно включаться только один из двух образующихся триозофосфатов, а именно – глицеральдегид-3-фосфат. Вследствие этого по мере потребления в ходе дальнейших превращений альдегидной формы фосфотриозы, дигидроксиацетонфосфат превращается в глицеральдегид-3-фосфат, т.е. дальнейшему окислению подвергается 2 молекулы фосфотриозы.
Образованием глицеральдегид-3-фосфата завершается первая стадия гликолиза.
Вторая стадия включает в себя окислительно-восстановительную реакцию, сопряженную с субстратным фосфорилированием, в процессе которого образуется АТФ.
В результате шестой реакции глицеральдегид-3-фосфат в присутствии фермента глицеральдегидфосфатдегидрогеназы (класс оксидоредуктазы), кофермента НАД+ и неорганического фосфата подвергается своеобразному окислению с образованием 1,3-бисфосфоглицериновой кислоты и восстановленной формы НАДН:
 1,3-бисфосфоглицерат представляет собой высокоэнергетическое соединение.
1,3-бисфосфоглицерат представляет собой высокоэнергетическое соединение.
Седьмая реакция катализируется фосфоглицераткиназой (класс трансферазы), при этом происходит передача богатого энергией фосфатного остатка на АДФ с образованием АТФ и 3-фосфоглицериновой кислоты:
 Данная реакция является реакцией субстратного фосфорилирования.
Данная реакция является реакцией субстратного фосфорилирования.
Восьмая реакция сопровождается внутримолекулярным переносом оставшейся фосфатной группы под действием фермента фосфоглицеромутазы (класс изомеразы).

Девятая реакция катализируется ферментом енолазой (класс лиазы):

Десятая реакция характеризуется разрывом высокоэнергетической связи и переносом фосфатного остатка от фосфоенолпировиноградной кислоты на АДФ (субстратное фосфорилирование).
Катализируется ферментом пируваткиназой (класс трансферазы):

Далее пути аэробного и анаэробного окисления расходятся.
В анаэробных условиях дальнейшие превращения пирувата приводят к образованию продуктов брожения.
Дата добавления: 2018-05-02; просмотров: 905; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!