ИЛИ ЦИТОПЛАЗМАТИЧЕСКОЕ, НАСЛЕДОВАНИЕ
Пластидное наследование. Пластиды представляют собой своеобразные лаборатории синтеза углеводов в растительном организме (см. гл.. 1). В ряде случаев установлено, что пластиды размножаются делением и расходятся в дочерние клетки во время их деления. Найдены примеры (род Cylindrocystis),когда в ходе оплодотворения сливаются две .клетки, каждая из которых несет по две пластиды, в результате чего образуется зигота с четырьмя пластидами. После мейоза каждая из гаплоидных клеток получает по одной пластиде, которая делится еще один раз. В результате образующаяся гамета содержит две пластиды. Таким образом, в мейозе происходит своеобразная редукция числа пластид.
Клетки, полностью утратившие пластиды, не способны их образовывать заново. Так, одноклеточная водоросль эвглена (Euglena mesmily) имеет 70—100 хлоропластов, но если ее содержать в темноте, то репродукция хлоропластов затормозится и по мере деления клеток могут возникнуть особи, совершенно лишенные пластид и неспособные образовать их заново.
Способность пластид быть носителями наследственных задатков была установлена уже давно. Совокупность пластид клетки как структур, передающих наследственную информацию, была названа пластидомом. Из всех структурных элементов цитоплазмы растений, с которыми можно связывать передачу потомству признаков материнского организма, пластиды наиболее удобны для анализа, так как в большинстве случаев они являются четко различимыми структурами, обладающими целым рядом морфологических особенностей.
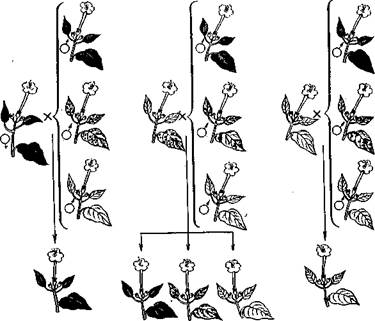
О первых фактах пластидного наследования сообщили Э. Баур и К. Корренс еще на заре развития генетики (в 1908 г.). Так, Корренс изучил наследование белой пестролистности у ночной красавицы (Mirabilis jalapa). У этого вида встречаются пестролистные растения, которые имеют в точках роста разные группы клеток: с пластидами, неспособными к образованию хлорофилла, и с нормальными пластидами. Вследствие этого иногда на растении образуются чисто зеленые или совершенно белые ветви. Семена, полученные с белых ветвей, дают нежизнеспособные всходы, так как у них не идет процесс фотосинтеза. При опылении цветков с пестролистных ветвей пыльцой от цветков с зеленых ветвей и при реципрокном скрещивании
171
 результаты получаются различными. В первом случае (? пестролистное X & зеленое) гибридные растения развиваются пестролистными, зелеными или белыми (гибнут). При реципрокном скрещивании (9 зеленое X с? пестролистное) в потомстве все растения оказываются зелеными. Цветки с зеленых ветвей дают только зеленое потомство, а с белых — белое (нежизнеспособное), независимо от того, пыльцой с каких растений они
результаты получаются различными. В первом случае (? пестролистное X & зеленое) гибридные растения развиваются пестролистными, зелеными или белыми (гибнут). При реципрокном скрещивании (9 зеленое X с? пестролистное) в потомстве все растения оказываются зелеными. Цветки с зеленых ветвей дают только зеленое потомство, а с белых — белое (нежизнеспособное), независимо от того, пыльцой с каких растений они
Развитие белых или зеленых частей растений из зиготы, содержащей пластиды обоих типов, определяется скоростью воспроизведения разных пластид и их распределением в ходе клеточных делений. Клетки, получившие только зеленые пластиды, дают зеленые участки тканей, а из клеток, имеющих только неокрашенные пластиды, образуются белые участки. Если клетка, имеющая два сорта пластид, разделится по линии АБ (рис. 76),
|

| 75. Наследование пестролистно-сти у Mirabilis jalapa. В качестве материнской формы взяты растения с листьями: / — зелеными, 2 — пестрыми, 3 — белыми. |
опылялись (рис. 75). На основании этого был сделан вывод, что наследование пестролистное™ связано с передачей и распределением при клеточных делениях двух типов пластид— зеленых и неокрашенных, причем передаются пластиды яйцеклеткой, в результате чего наследование осуществляется по материнской линии. Однако в редких случаях, например у герани (Pelargonium zonale), пластиды передаются не .только яйцеклеткой, но и спермием, содержащим цитоплазму. При этом пестролистность наследуется не только по материнской, но и по отцовской линии, 172

то образуются две клетки, которые 76.
| Схема случайного распределения белых и зеленых пластид при клеточном делении: /— образование двух клеток, из которых одна (а) даст белы* участок, а другая (б) — пестрый; 2—образование двух клеток, из которых одна (в) даст зеленый участок, а другая (г) — пестрый, АБ и ВГ — линии деления клеток. |
дадут два участка (белый — а и пестрый—б), при разделении по линии ВГ — зеленые и пестрые (в, г).
У некоторых растений наблюдается закономерная изменчивость проявления пестролистности в ходе онтогенеза. Например, у пестролистного хмеля (Humulus Japonica) семядоли всегда бывают чисто зелеными, а на листьях пестролистность тем сильнее, чем позднее они формируются.
У подорожника (Plantago) развитие пестролистности в значительной степени зависит от температуры. Так, при низких температурах пестролистность значительно сильнее, а повышение температуры приводит к ее ослаблению или даже полному исчезновению. Возможно, что это связано с различной скоростью размножения разных пластид.
Установлено, что свойства пластид часто определяются непосредственно ядерными генами и наследование пестролистности при этом осуществляется в соответствии с установленными закономерностями ядерной наследственности, но измененные
473

|
пластиды передаются вместе с цитоплазмой. Такие случаи известны у ячменя, кукурузы и других растений.
Наследование через митохондрии. Митохондрии имеют непосредственное отношение к процессам дыхания (см. гл. 1). Считается, что они способны к самовоспроизведению посредством поперечного деления. В них обнаружена ДНК. В результате деления дочерние клетки получают от материнской примерно половину ее митохондрий.
У некоторых грибов (дрожжи, нейроспора) была обнаружена дыхательная недостаточность, которая обусловлена необратимыми наследственными изменениями функции митохондрий— у них утрачена активность цитохромоксидазы.
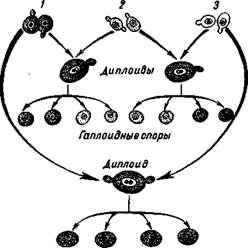
Б. Эфрусси обнаружил штаммы дрожжей Saccharomyces cerevisiae, которые спонтанно образуют карликовые колонии с дыхательной недостаточностью. Поскольку они возникают при вегетативном размножении гаплоидных дрожжей, они были названы вегетативными карликовыми. Наряду с вегетативными карликовыми колониями была обнаружена форма, по фенотипу — росту и дыхательной недостаточности — сходная с первой, но она давала расщепление по признаку карликовости, как будто он определялся одним ядерным геном. Эта форма была названа расщепляющимся карликовым штаммом.
Так как у дрожжей при скрещивании и образовании зиготы сливаются целые клетки, то в этом случае можно оценить роль цитоплазмы и ядра в наследовании при скрещивании таких карликов с нормальной формой. Генетический анализ вегетативного и расщепляющегося карликовых штаммов (рис. 77) показывает, что фенотип расщепляющейся карликовости определяется ядерным геном, поскольку при скрещиваниях наблюдается расщепление аскоспор в отношении 1:1. При скрещивании вегетативных карликов и нормальных дрожжей дицлоидная зигота, в которой есть митохондрии от нормальной формы, не дает расщепления — из аскоспор не появляются мелкие колонии. Следовательно, у этих форм геномы одинаковы, различалась лишь цитоплазма. Расщепления же по типу цитоплазмы в мейозе не происходит. В данном эксперименте факт цитоплазм этического наследования очевиден.
Получено и прямое доказательство роли митохондрий в наследственной передаче дыхательной недостаточности у дрожжей. Вегетативных карликов, лишенных клеточных оболочек, выращивали в присутствии изолированных митохондрий нормальных дрожжей. В результате часть образовавшихся колоний (2—2,5%) имели нормальные размеры. Этот факт можно объяснить, предположив, что «нормальные» митохондрии, попав в клетки вегетативных карликов, исправили дефект их дыхательной системы и, передаваясь из клетки в клетку в ходе деления, способствовали образованию нормальных колоний.
174
77.
Схема генетического анали
за вегетативных и расщеп
ляющихся карликовых
штаммов у дрожжей:
/ — нормальный штамм; карликовые штаммы; 2 — расщепляющийся, 3 — вегетативный.
Гаплоидные споры
Цитоплазматическая мужская стерильность. В некоторых случаях наблюдаются все закономерности цитоплазматического наследования, но не удается вычленить определенные структуры, ответственные за наследование соответствующего признака.
Один из самых ярких примеров этого — явление цитоплазма- тической мужской стерильности (ЦМС), обнаруженное у многих растений — кукурузы, лука, свеклы, льна и др. Цитоплазматическая мужская стерильность у кукурузы была открыта в 30-х годах одновременно в СССР М. И. Хаджиновым и в США М. Род-сом. Кукуруза—-однодомное растение, женские цветки у нее собраны в початок, мужские — в метелку. У некоторых сортов кукурузы были обнаружены растения, имевшие в метелках недоразвитые пыльники, часто совершенно пустые, а иногда с недоразвитой стерильной пыльцой. Оказалось, что этот признак определяется особенностями цитоплазмы. Опыление растений с мужской стерильностью нормальной пыльцой с других растений в большинстве случаев дает в потомстве растения со стерильной пыльцой. При повторении этого скрещивания в течение ряда поколений признак мужской стерильности не исчезает, передаваясь по материнской линии. Даже тогда, когда все 10 пар хромосом растений со стерильной пыльцой замещаются хромосомами от растений с фертильной пыльцой, мужская стерильность сохраняется. Это послужило убедительным доказательством того, что наследование данного признака осуществляется через цитоплазму. Цитоплазма, обусловливающая стерильность
175
|
|

|
|

|
|

|
|

| u,ums rfrf uumN RfRf |
| Стерильность |
| Фертильность |
| uums Rfrf |
| uumN rfrf |
| Фертильность uums Rfrf |
| Фертильность |
78.
| uums rfrf |
|
|

|
|

Схема наследования цито-
плазматической мужской
стерильности. Цитоплазма
цитв — стерильная, цитИ —
нормальная Rf — ген-восста
новитель фертильности
пыльцы.
| Стерильность |
Фертильность
пыльцы, была обозначена символом uuts (стерильная цитоплазма), а цитоплазма растений с фертильной пыльцой — символом цитп (нормальная цитоплазма).
Установлено, что генотип растения может оказывать определенное влияние на действие стерильной цитоплазмы. Цитоплазма цит* может обусловить стерильность пыльцы только при наличии в генотипе растения рецессивного гена rf в гомозиготном состоянии rfrf. Если же этот ген представлен доминантной аллелью Rf, то растение цитв RfRf или цитв Rfrf имеет нормальную пыльцу. Аллель Rf является, таким образом, восстановителем фертильности пыльцы. Следовательно, фертильную пыльцу могут иметь и растения цит1* rfrf, и uutn RfRf или Rfrf, и цитв RfRf или Rfrf, а полностью стерильную — только растения цитБ rfrf. Многократное повторение скрещивания 9 цитв rfrfX с? uutn rfrf всегда дает потомство с полностью стерильной пыльцой. И только в случае скрещивания uuts rfrfXt{UTs RfRf (или quTN RfRf) может быть получено потомство, где все растения будут иметь нормальную пыльцу, несмотря на наличие цитоплазмы uuts (рис. 78). Следует еще раз подчеркнуть, что ген Rf не изменяет структуру и специфичность цитоплазмы цитв, а лишь тормозит проявление ее действия.
Исследование цитоплазматической мужской стерильности представляет пример успешного генетического анализа ядерно-цитоплазматических отношений.
3. ПРЕДЕТЕРМИНАЦИЯ ЦИТОПЛАЗМЫ
Сущность предетерминации цитоплазмы. В ряде случаев наследование признаков связано с особенностями цитоплазмы, причем эти особенности могут возникать в процессе индивидуального развития организма либо под влиянием факторов внешней среды (онтогенетическая или фенотипическая предетермина- ция), либо под влиянием генотипа (генотипическая предетерми- нация).
Онтогенетическая предетерминация. Известны факты, когда наследование некоторых признаков по материнской линии обусловлено изменениями в цитоплазме, возникающими в ней под влиянием определенных внешних факторов. Обычно такие изменения, появившиеся в одном поколении, нестойки и через несколько поколений постепенно исчезают, возвращаясь к исходному типу.
Например, воздействие повышенной температурой на яйца самок наездника (Habrobracon juglandis) до оплодотворения приводит к изменению окраски тела у их потомства. В последующих поколениях при размножении в нормальных температурных условиях это изменение постепенно затухает. Когда же температурному воздействию подвергаются самцы, а самки выращиваются в нормальных условиях, подобного эффекта не наблюдается.
Подобные изменения, затухающие в ряду поколений при возвращении организмов в исходные условия, называются длительными модификациями.
В основе большей части случаев онтогенетической предетерминации, по-видимому, лежит вырабатывающееся под воздействием различных факторов внешней среды приспособление яйцеклеток, которое передается потомству по материнской линии при половом размножении и через соматические клетки в случае бесполого размножения. Механизм такой передачи до сих пор не выяснен.
Длительные модификации могут постоянно сохраняться в ряду поколений при условии сохранения вызывавших их факторов; при отсутствии последних происходит постепенный возврат к исходному состоянию (см. гл. 23).
Генотипическая предетерминация. Еще больший интерес представляют факты предетерминации свойств цитоплазмы под влиянием генотипа материнского организма. Ярким примером генотипической предетерминации цитоплазмы является наследование направления завитка раковины у некоторых пресноводных моллюсков (Limnaea), которые являются гермафродитами и могут размножаться как путем самооплодотворения, так и путем скрещивания. Встречаются два типа закручивания раковины: против часовой стрелки — левозакрученные и по ходу
176
7 Генетика с основами селекции
177

79. часовой стрелки —правозакрученные.
| Роль цитоплазмы в наследовании направления завитка раковины у моллюска Limnaea: D — ген, определяющий право-закрученность, d — левозакру-ченность. |
Направление закручивания раковины определяется одной парой аллелей: правозакрученность D доминирует над левозакрученностью d.
При реципрокных скрещиваниях гибриды Fu имеющие один и тот же генотип Dd, различаются по фенотипу. В скрещивании 9 DDx<? dd все гибридные особи имеют материнский тип — правозакрученные раковины. В скрещивании ? ddX Xе? DD потомство также имеет материнский тип завитка, т. е. левозакрученную раковину (рис. 79).
От самооплодотворения гетерозиготных форм F\ {Dd) в обоих скрещиваниях все потомки F2 обладают правозакрученной раковиной, хотя гибриды F\ (материнские формы) различались по фенотипу. Когда было исследовано потомство от каждой особи в отдельности, то выяснилось, что 'Д семей имели левый завиток, а 3Д — правый. Простое менделевское расщепление по данной паре признаков 3: 1 выявилось не в F2, а только в F3. При этом типе наследования фенотип потомков соответствует генотипу матери, а не генотипу зигот, из которых они развиваются. Это может быть только при допущении, что данный признак предопределяется генотипом материнского организма в цитоплазме яйца в процессе его развития. Рассмотренный тип наследования и является в собственном смысле слова материнским.
178
Как было выяснено, направление завитка раковины определяется характером спирального дробления оплодотворенного яйца, т. е. расположением бластомеров по спирали вправо или влево, что в свою очередь зависит от ориентации веретена при втором делении дробления.
Рассмотренный пример, строго говоря, не относится непосредственно к цитоплазматическому наследованию, поскольку здесь свойства цитоплазмы детерминированы действием хромосомных генов, а не элементами самой цитоплазмы, т. е. здесь действует механизм хромосомного наследования, который изменяет цитоплазму яйцеклетки еще до оплодотворения.
Н. Н. Соколов провел исследования генотипической преде-терминации у реципрокных гибридов между видами дрозофилы — Drosophila virilis и D. littoralis. Гибриды от скрещивания D. littoralisXD. virilis развивались нормально, а при обратном скрещивании почти все потомки обнаружили сильное нарушение развития и появление различных аномалий в строении крыльев, глаз, ног и т. д.
Цитологический анализ гибридов от второго скрещивания показал, что у них наблюдаются различные нарушения в наборе хромосом и патологические митозы в соматических клетках; Атипическое деление клеток было обнаружено на ранних стадиях развития гибридных зародышей, чем и объясняются, по-видимому, уродства и аномалии у имаго.
Так как нарушения митоза не наблюдались у гибридов от первого скрещивания, то причину нарушений нужно искать в особенности цитоплазмы D. virilis. Однако в F2 гибриды обоих типов имели небольшое количество атипических митозов. Оказалось, что видовые различия цитоплазмы, проявившиеся у гибридов Fu в F2 исчезли. Сходство гибридов F2 автор объясняет тем, что они происходят из яйцеклеток, сформировавшихся в самках Fu которые имели одинаковый генотип в случае обоих реципрокных скрещиваний.
Чтобы выяснить роль ядра в предетерминации свойств цитоплазмы яйцеклетки, Соколов провел дополнительную серию скрещиваний. Гибрид D. littoralisXD. virilis, нормально^ развившийся в Fu повторно скрещивался с отцовской формой. После полного замещения у этого гибрида хромосом материнского вида хромосомами отцовского, достигнутого к четвертому поколению возвратных скрещиваний, была получена новая форма, самки которой при скрещивании с самцами D. littoralis давали в потомстве столь же высокий процент атипических митозов, как и гибриды D. virilis XD. littoralis. В результате беккроссов цитоплазма полностью изменилась в сторону свойств цитоплазмы D. virilis под влиянием хромосом этого вида. Следовательно, именно генотипическая предетерминация определяет несовместимость цитоплазмы одного вида с хромосомами другого.
| 7* |
179
4. НАСЛЕДОВАНИЕ ЧЕРЕЗ ИНФЕКЦИЮ И ВКЛЮЧЕНИЯ
Наследование через инфекцию. Через цитоплазму могут передаваться различные субмикроскопические частицы и симбионты клетки, которые обладают свойством саморепродуцироваться и в силу этого способны имитировать цитоплазматиче-ское наследование.
У мышей имеется линия с наследственной предрасположенностью к развитию рака молочной железы. При этом предрасположение к заболеванию передается по материнской линии и только при выкармливании потомства. Если к матерям-кормилицам из раковых линий подсадить мышат из нераковой линии, то такие мышата также становятся предрасположенными к раку молочной железы. Если мышат из раковой линии с момента рождения вскармливают нормальные кормилицы, то мышата остаются здоровыми. Таким образом, опухоли в данном случае вызываются инфекцией через молоко матери. Этот инфекционный агент и был назван фактором молока. Установлено, что он имеет вирусную природу.
Другим примером наследования через инфекцию цитоплазмы являются линии дрозофилы, в которых отсутствуют самцы,— так называемые бессамцовые линии. Самки, несущие фактор бес-самцовости, в потомстве дают только женский пол, так как самцы гибнут на эмбриональной стадии. Фактор бессамцовости может быть передан нормальным самкам путем инъекции цитоплазмы, взятой из неразвившихся яиц бессамцовой линии. Инъекция такой цитоплазмы взрослым самцам вызывает их гибель. Оказалось, что фактор бессамцовости представляет собой спирохету, которая избирательно размножается в цитоплазме половых клеток самок и губительна для соматических клеток самцов.
Наследование через включения. У инфузорий Paramecium aurelia известны линии, которые выделяют в среду специфическое вещество, называемое парамецином. Сами продуценты па-рамецина не страдают от него, но парамеции из других линий от этого вещества гибнут. Инфузории, выделяющие парамеции, были названы «убийцами». Установлено, что они содержат в своей цитоплазме особые частицы, по виду напоминающие риккетсии, в состав которых входит ДНК; они были названы каппа-частицами. Только имея эти частицы, инфузория может продуцировать парамеции. Чувствительные к парамецину инфузории этих частиц не содержат.
Сохранение каппа-частиц в цитоплазме и выделение параме-цина инфузорией-«убийцей» контролируется доминантным геном К; его рецессивная аллель k не способствует их сохранению. При прямом делении инф.узории-«убийцы» постоянно дают однотипный клон со свойствами «убийц». При соответствующих
180
условиях опыта удается произвести скрещивание, т. е. вызвать конъюгацию двух клеток — «убийцы» и чувствительной к парамецину.
На рисунке 80 представлена схема наследования аллелей К и А и распределения каппа-частиц. Исходные формы являются гомозиготными (КК и kk) , эксконъюганты — гетерозиготными (Kk). При последующей автогамии эксконъюгантов происходят два деления созревания микронуклеуса. Из возникших четырех гаплоидных ядер три дегенерируют, а оставшееся делится мито-тически. В результате образуются два гаплоидных пронуклеуса, которые затем сливаются, давая в конечном итоге диплоидный и обязательно гомозиготный микронуклеус (в рассматриваемом примере КК или kk). В результате автогамии эксконъюгантов будет наблюдаться расщепление по генотипу в соотношении 1 : 1 «убийц» (КК) к чувствительным (kk), но распределение каппа-частиц будет зависеть от длительности конъюгации родительских клеток. Если конъюгация была кратковременна и происходил обмен лишь микронуклеусами, а цитоплазмой эксконъюганты не успели обменяться, то каппа-частицы не попадут в цитоплазму чувствительного партнера и останутся только в исходной клетке. В том случае, если конъюгация была достаточно продолжительной, то происходящий от чувствительной клетки эксконъюгант получит не только ген К и станет по генотипу Kk, но и цитоплазму с каппа-частицами. При последующем делении эта парамеция даст клон «убийц». Каппа-частицы будут размножаться в цитоплазме инфузории и сохраняться при наличии аллели К- Если же они попадут в цитоплазму чувствительной клетки (kk), то они не размножаются и в ряду делений как бы постепенно «разбавляются», впоследствии исчезая. Предполагают, что эти частицы являются симбионтами клетки.
Все разобранные нами случаи показывают, что в цитоплазме иногда обнаруживаются различные включения, или частицы, способные к самовоспроизведению, которые могут быть передатчиками ряда свойств по материнской линии. Но эти частицы по существу не являются неотъемлемыми элементами живой клетки. К подобным случаям следует относить также передачу через цитоплазму какого-либо инфекционного начала — вирусов или бактерий.
ГЕНЕТИЧЕСКИЙ АНАЛИЗ
НЕХРОМОСОМНОГО НАСЛЕДОВАНИЯ
Генетическая система клетки. Изучение нехромосомного наследования представляет существенный интерес для выяснения генетической системы клетки в целом.
До сих пор под термином генотип мы имели в виду лишь систему генов, локализованных в хромосомах, Наследственные
181
X |

К о н ъ ю га и. и я
/ \
|
|

| Л А Автогамия |
 короткая, длинная
короткая, длинная
/ \
Л /\ Л Л Л Л /\ Л
АДЛЛЛЛЛЛ ЛЛЛЛАЛЛЛ
80.
Схема наследования аллелей гена К и каппа-частиц у инфузорий. Каппа-частицы изображены черными точками.
факторы, локализованные в цитоплазме и ее органоидах, обозначаются термином плазмотип, или плазмон. Правильнее было бы расширить понятие генотипа и включить в него как систему хромосомных генов (геном), так и цитоплазматических (плазмон). Любая единица цитоплазматической наследственности, соответствующая единице хромосомной наследственности (гену), называется плазмогеном (см, схему).
Дата добавления: 2020-01-07; просмотров: 426; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!