Транспорт вирусный частиц по клетке
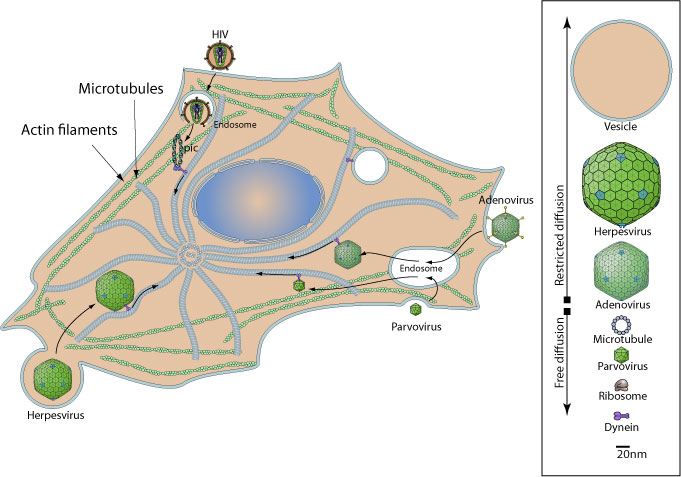
Третий этап — внутриклеточные перемещения вирусов.
После интернализации вирусаи его «раздевания» генетический аппарат целенаправленноперемещается в цитоплазме по цитоскелету, заимствуя клеточные молекулярные моторы. Диффузионное перемещение надмолекулярных геномных комплексов вируса в переполненной цитоплазме маловероятно, так как оно возможно лишь для структур меньше 20 нм (соответственно менее 500 кДа).
В вирусном перемещении участвуют как миозиновый мотор с актиновой микросетью, так и динеин-актиновый мотор, ассоциированный с микротрубочковыми путями. Микротубулярный цитоскелет связан с актиновым кортексом, поэтому внутриклеточный путь у некоторых вирусов начинается при взаимодействии с актиновыми филаментами.
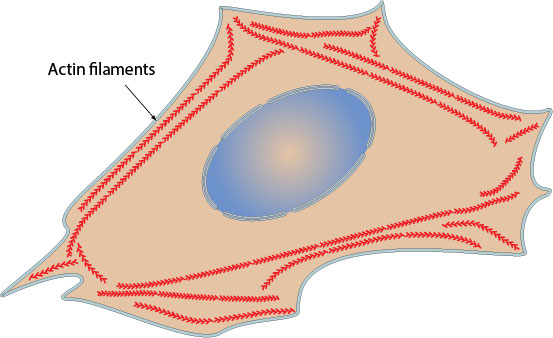
Актин-зависимый транспорт вызывается вирусными белками, которые взаимодействуют с актином и/или «хозяйскими» клеточными моторными белками, такими как миозин или белками, способствующими полимеризации/ деполимеризации актина.
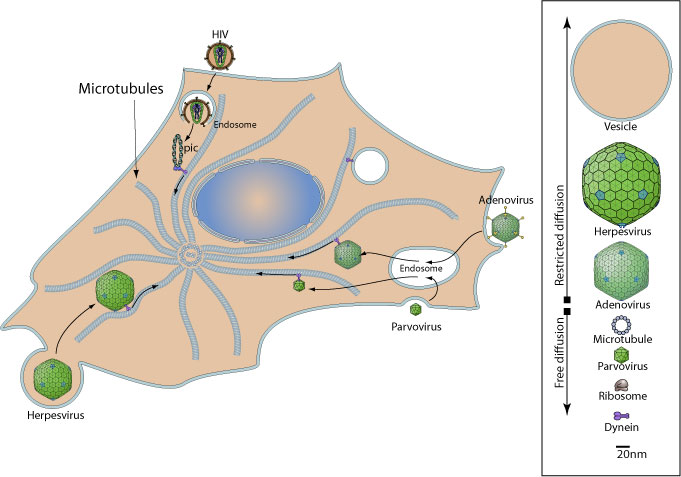 Микротрубочко-зависимый транспорт вовлекает моторные белки, такие как динеин и кинезин, и в основном используются вирусами, «нацелившими» свой геном в ядра.
Микротрубочко-зависимый транспорт вовлекает моторные белки, такие как динеин и кинезин, и в основном используются вирусами, «нацелившими» свой геном в ядра.
С помощью микротрубочек
ü Adenoviadae (Ad5)
ü Herpesviridae(HHV-l)
ü Rhabdoviridae (вирусбешенства)
ü Retroviridae (HIV-1)
ü Hepadnaviridae (HBV)
ü Papillomaviridae (HPV-16)
С помощью активного цитоскелета:
Picornaavindae (Poliovirus)
Вход и выход вирусов из ядра. Варианты выхода вирусов из клетки. Вирусные фабрики.
Вход и выход вирусов из ядра
Вход
В ядерном транспорте вирусом используется клеточный аппарат т.н. «комплекса ядерных пор», движение в котором регулируется более чем 50-клеточными белками-нуклеопоринами, а верхний пропускной лимит составляет 39 нм.
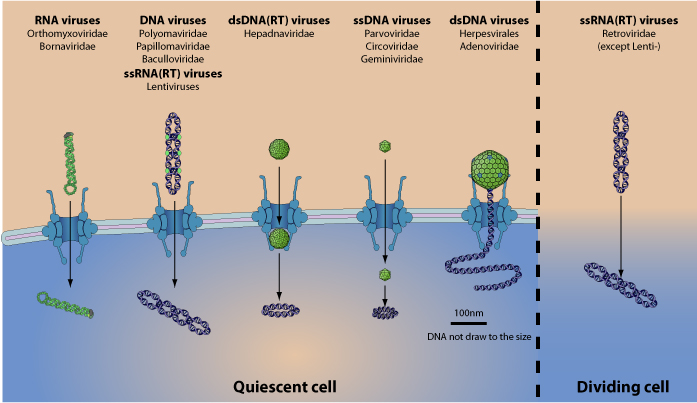
Среди вирусов существует заметное разнообразие в механизмах внутриядерного транспорта.
Мелкие нуклеокапсиды парвовирусов (20 нм) и вируса гепатита В (38 нм) проходят в ядра в интактной форме и уже дополнительно дезинтегрируются внутри ядер.
Нитевидные нуклеокапсиды вируса гриппа А (20 нм) взаимодействуют с импортином (3 и проходят в ядра клеток в интактной форме.
Более крупные вирусы, такие как аденовирус 2, последовательно связывают белок NUP214/С AN экстрануклеарных филаментов и затем кариоферины (3 и 7 и гистона Н1), что вызывает диассамблирование крупного нуклеокапсида на входе в ядерную пору, и уже свободная вгДНК входит в ядро.
Нуклеокапсид вируса герпеса (120 нм) «причаливает» к ядерной поре через импортин β и свободная вгДНК впрыскивается в ядро без распада нуклеокапсида.
Внутриядерный транспорт прединтегразного комплекса лентивируса ВИЧ-1 не требует ядерных пор и осуществляется на стадии митоза клеток, когда ядерная оболочка временно отсутствует, и контролируется вирусными белками МА, интегразой, vpr и элементом flap гДНК
Выход
Все ядерно-реплициирующие вирусы должны получить свое «потомство» из ядра. Этот процесс требует особых вирусных белков, т.к. этот процесс является не естественным для клетки, когда ДНК или несшитые РНК материал покидают клеточное ядро.
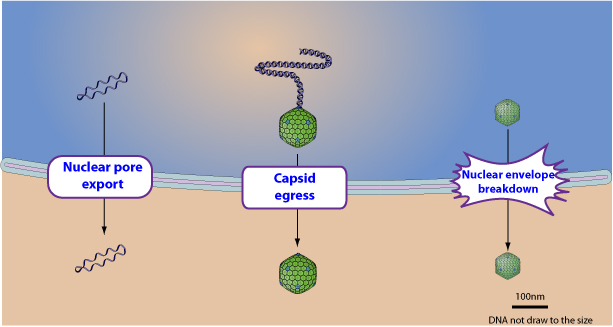
Три основных способа позволяют ядерным вирусам покидать эту органеллу:
- Ядерный поровый экспорт: экспорт генетического материала с белком-шаппероном;
- Ядерный выход: слияние с ядерной мембраной, чтобы выйти в цитоплазму;
- Распад ядерной оболочки.
Ядерный поровый экспорт

(зеленькие шарики – белки-шаппероны)
Ядерный выход
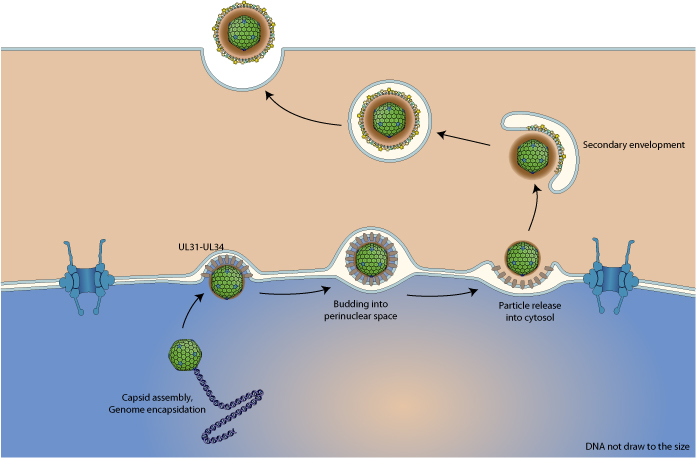
Вирусные капсиды, собранные в ядре, могут мигрировать в цитоплазму за счет использования необычного транспорта – ядерного выхода. В этом процессе вирусный капсид подается в перинуклеарное пространство, производящий вспомагательную визикуляцию после фузии. После слияния голый капсид освобождается в цитозоле.
Дата добавления: 2018-06-01; просмотров: 1367; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!