Молекулярный механизм кроссин-
говера. Представление о нуклеиновых
кислотах как носителях наследственной информации позволило выдвинуть ряд гипотез, объясняющих и механизм рекомбинации сцепленных генов, т. е. механизм кроссинговера. Различные варианты этих гипотез сводятся к идее, сформулированной в 1963 г. К. Уайтхаузом. В основу предполагаемого механизма положено наиболее характерное свойство молекулы ДНК, а именно способность образовывать водородные связи между комплементарными основаниями А—Т и Г—Ц. Рассмотрим гипотезу в общей форме (рис. 119).
Весь процесс обмена участками двух гомологичных хромосом, считая, что каждая хроматида представлена одной молекулой ДНК, можно разбить на несколько этапов: разрыв одной цепи в близких, но не идентичных точках обеих молекул ДНК (рис. 119, 2); разрыв водородных связей между комплементарными цепочками ДНК вблизи разрывов и образование новой


| 119. Возможный механизм рекомбинации на молекулярном уровне (стрелками указаны точки первоначальных разрывов). Сплошные линии— одинарные цепочки спиралей ДНК; пунктирные— вновь синтезированные цепочки ДНК; 1—8 — этапы рекомбинации. |
биспиральной «гибридной» структуры молекулы ДНК (рис. 119,3), локальный синтез ДНК на месте отошедших цепочек (рис. 119,4), расхождение вновь синтезированных цепочек в стороны (рис. 119,5) и их комплементарное соединение (рис. 119,6), разрыв цепочек, не претерпевших рекомбинацию (рис. 119,7), и замыкание разорванных концов (рис. 119,5).
Все этапы процесса происходят в ранней профазе мейоза, т. е. уже после того, как закончилась редупликация хромосом. Предложенный механизм рекомбинации удовлетворительно объясняет большинство экспериментальных фактов, полученных на различных генетических объектах.
|
|

| и БУ |
Г
и'
118.
Замещение пар.ос-
нований: А—Т на
Г—Ц и Г—Ц
на А—Т.
Пунктирная стрелка указывает на ошибочное спаривание, число точечных линий между нуклеотн-дами соответствует числу водородных связей.
Таким образом, очевидно, что ген имеет сложное строение, он представляет собой единицу, далее делимую при кроссинго-вере, и не представляет собой единицы мутирования. Однако как молекулы белка нельзя разделить на части без потери ее специализации, так и ген как единица наследственной информации остается функционально неделимым.
286

|

Раздел V. Генетика пола
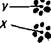
Гинандроморф дрозофилы.
Вверху — предполагаемая схема возникновения; X — элиминируется.
Проблема пола, т. е. вопрос о механизмах, которые определяют развитие мужских и женских особей, возникла еще в глубокой древности. Причину этого надо искать в ее практическом значении. Однако до сих пор эта проблема остается одной из актуальных и еще не решена окончательно. Решение ее имеет не только практический интерес, но и важное теоретическое значение. Оно позволит вскрыть генетические механизмы определения и становления одного из сложных признаков организма в онтогенезе, а также будет способствовать выяснению вопроса о соотношении дискретного определения признаков и свойств, с одной стороны, и целостности организма и генотипа — с другой.
Глава 18. ОПРЕДЕЛЕНИЕ ПОЛА
Пол— это совокупность признаков и свойств организма, обеспечивающих его участие в воспроизводстве потомства и передаче наследственной информации за счет образования гамет.
Прежде чем излагать генетические основы определения пола, необходимо познакомиться с биологией пола.
БИОЛОГИЯ ПОЛА
Обычно признаки, по которым отличаются особи разных полов, делят на первичные и вторичные половые признаки. К первичным относят те морфологические и физиологические особенности организма, которые обеспечивают образование гамет и соединение их в процессе оплодотворения. К числу их относятся, например, гонады, половые пути и наружные гениталии у высших животных, андроцей и гинецей у высших растений. К вторичным половым признакам относят признаки и свойства организма, непосредственно не обеспечивающие процессы гаметоге-неза, спаривания и оплодотворения, но играющие некоторую вспомогательную роль в половом размножении. К ним относятся особенности строения плавников у рыб, оперения у птиц, грудных желез у млекопитающих, длины междоузлий и сроков цветения у высших растений и др. Из числа вторично-половых признаков выделяют иногда ограниченные полом признаки. Генетическую информацию о них несут все особи, но проявляются они только у одного пола. Так, быки несут гены, определяющие молочность, петухи — гены, определяющие яйценоскость, но действие их у самцов не проявляется.
Следует отличать также признаки, зависимые от пола. К числу зависимых от пола признаков относятся такие признаки, характер доминирования которых в гетерозиготе зависит от пола особи. Так, доминантные гомозиготные овцы (НН) рогаты, а гомозиготные рецессивы (hh) безроги, независимо от пола. Однако гетерозиготные (Hh) самцы рогатые, а самки безрогие. Аналогично наследуется раннее облысение у человека. Доминирование в таких случаях определяется количеством мужских и женских половых гормонов в крови. Знание закономерности наследования признаков, зависимых от пола, позволяет еще глубже понять сущность явления доминирования и подойти к практическому управлению доминированием.
Различия особей мужского и женского пола — половой диморфизм встречается у некоторых низших и многих высших растений и животных. Среди высших растений около 5000 видов двудомны. У высших животных обоеполые (гермафродитные) формы в норме не встречаются. Во всех случаях отличия касаются морфологических, биохимических, физиологических признаков, экологии, поведения у животных и т. д. Однако'на низших ступенях эволюции у одноклеточных организмов отличия особей, вступающих в копуляцию, могут быть невидимы и касаться лишь физиологических особенностей, поэтому и пол их обозначают условно как « + пол» или «—пол». Различия между этими двумя типами часто носят количественный характер, так что у многих одноклеточных водорослей (хламидомонады, например), некоторых низших грибов или инфузорий удается выделить несколько линий, внутри которых копуляция клеток невозможна, а между линиями она осуществляется. Удалось сгруппировать многочисленные линии в « + » и «—» типы, но оказалось, что и внутри типа копуляция клеток разных линий иногда происходит.
Следовательно, здесь можно говорить об относительной сек суальности.
Относительная сексуальность особенно хорошо изучена у хламидомонад М. Гартманом и Ф. Мевусом. Они показали, что гаметы даже одного пола, но с разной половой валентностью способны воспроизводить потомство половым путем. Половые различия гамет связаны с гормоноподобными веществами (га-монами).
В целом можно сказать, что половой диморфизм очень широко распространен в природе и встречается на всех ступенях эволюции.
Почему же возник половой диморфизм? Для ответа на этот вопрос следует вспомнить, что биологический смысл полового процесса состоит в перекомбинации наследственной информации,, ведущей к увеличению общего фонда наследственной изменчивости. Впервые на это обратил внимание Ч. Дарвин. Он сформулировал закон биологической полезности скрещивания и объяснил им происхождение двух полов.
Как уже было сказано, проблема пола — старая проблема. Первые попытки ее разрешения были сделаны еще Аристотелем. Было высказано много сотен различных гипотез, но они не только не решали проблему, но делали ее одной из самых запутанных и мистических. Научное решение стало возможно лишь после развития генетики (открытие законов Менделя) и цитологии (изучение кариотипов).
290
2. ХРОМОСОМНАЯ ТЕОРИЯ ОПРЕДЕЛЕНИЯ ПОЛА
Сущность хромосомной теории определения пола. Очень
давно люди заметили, что соотношение полов у раздельнополых организмов близко к 1 : 1, т. е. самцы и самки встречаются одинаково часто. Ниже указан процент мужских особей у разных организмов.
Человек............................ 51 Мышь.......................................... 50
Лошадь............................. 52 Курица....................................... 49
Крупный рогатый скот 50—51 Утка........................................... 50
Овца.................................. 49 Голубь.................................... .50
Свинья.............................. 52 Дрема луговая........................... 49
Собака............................... 56 Конопля..................................... 45
Еще Мендель обратил внимание, что такое же расщепление 1 : 1 характерно для анализирующего скрещивания: АаХаа. Было высказано предположение, что один из полов должен быть гомозиготным, а другой — гетерозиготным. Первое экспериментальное доказательство в пользу этой гипотезы было получено К. Корренсом. Среди рода Bryonia (переступень) есть двудомные (В. dioica) и однодомные (В. alba) виды. Для того чтобы определить, как наследуют пол мужские и женские растения двудомного вида, было произведено скрещивание их с однодомным. Оказалось, что в потомстве женских растений были только женские, а в потомстве мужских — половина женских и половина мужских растений. Отсюда был сделан вывод, что женские растения Bryonia гомозиготны, а мужские — гетерозиготны.
Пол, образующий одинаковые в отношении определения пола гаметы, назвали гомогаметным, а пол, образующий разные гаметы,— гетерогаметным.
Решающее доказательство в пользу такого заключения, как было уже сказано (см. гл. 8), получили цитологи. Еще в конце прошлого века у клопа Lygaeus при изучении сперматогенеза были описаны гаплоидные сперматоциты II двух сортов: с ^-хромосомой и У-хромосомой, в отличие от самок, которые в яйцеклетках, кроме 6 аутосом, одинаковых с самцами, обязательно имели А-хромосому (рис. 120). У другого клопа Protenor гетерогаметным полом также оказался мужской. Но у этого вида половина сперматоцитов, кроме 6 аутосом, имела А-хромосому, а половина ее не имела (рис. 120).
Было высказано предположение, что А- и У-хромосомы имеют отношение к определению пола, их назвали половыми хромосомами. Экспериментальные доказательства этого были получены Т. Морганом и его сотрудниками при изучении наследования признаков, сцепленных с полом (см. гл. 8). Так была впервые сформулирована хромосомная теория определения пола.
Половые хромосомы и их роль в определении пола. Это открытие стимулировало дальнейшие цитологические исследова-
ния. Половые хромосомы были найдены у многих организмов. Среди растений впервые половые хромосомы были описаны у печеночного мха Sphaerocarpus. Известны они у высших растений: меландриума, щавеля, элодеи, хмеля и других. У животных они описаны для многих насекомых, птиц, млекопитающих. Описаны они и у человека.
|
|

 Изучение половых хромосом показало, что они отличаются от аутосом не только генетически (см. гл. 8), но и цитологически. Половые хромосомы богаты гетерохроматином (см. гл. 2). Редупликация их происходит асинхронно с аутосомами, а у го-могаметного пола одна из Х-хромосом репродуцируется позже
Изучение половых хромосом показало, что они отличаются от аутосом не только генетически (см. гл. 8), но и цитологически. Половые хромосомы богаты гетерохроматином (см. гл. 2). Редупликация их происходит асинхронно с аутосомами, а у го-могаметного пола одна из Х-хромосом репродуцируется позже
| Protenor betfraai |
|
|
| Lygaeus turcicus |
 Анафаза I Гаметы Анафаза I Гаметы
Анафаза I Гаметы Анафаза I Гаметы
| 120. |
| Кариотипы самцов и самок и хромосомные наборы гамет гетерогаметного пола. |
остальных. В мейозе они часто сильно спирализованы (гетеропикноз). Половые хромосомы у гетерогаметного пола (гетероморфные пары) не конъ-югируют или конъюгируют лишь частично, что указывает на гомологичность лишь отдельных участков. Как уже говорилось (см. гл. 8), при расхождении А- и У-хромосом в редукционном делении образуются 2 разные клетки: одна с А-хромосомой, другая — с У-хромосомой, следовательно, соотношение гамет с Х- и У-хромосомой, образуемых гетерогаметным полом, бывает точно 1:1. Точно так же два сорта гамет образуются, если клетка содержит одну А-хромосому, при этом 50% гамет имеет А'-хромосому, а 50% не имеет ее. Гаметы, образуемые гомогаметным полом, все одинаковые и содержат А'-хромосому (название гомогаметный и указывает на это). В результате оплодотворения возникает равное количество самцов и самок. Иными словами, хромосомный механизм определения пола является идеальным саморегулирующимся механизмом. Анализ половых хромосом у различных организмов показал, что существуют разные типы хромосомного определения пола (табл. 14). Они получили название тип ХО и тип ХУ. Гетерогаметным полом может быть как мужской, так и женский. Сейчас описаны и более сложные комплексы половых хромосом, но они принципиально не отличаются от только что названных.
292
293
Таблица 14
Типы определения пола
| Тип | Гетерогамет-ный пол | Гаметы | Зиготы | Организмы | ||
| Самцов | Самок | Самцов | Самок | |||
| ХУ ХО ХУ ХО | Мужской » Женский » | X и У X и 0 X X | X X X и У X и 0 | ХУ ХО XX XX | XX XX ХУ ХО | Человек, млекопитающие, дрозофила, мелан-дриум и др. Клоп (Protenor), кузнечики Dioscorea и др. Птицы, земноводные, рептилии, бабочки, клубника и др. Моль и др. |
Гинандроморфизм. Иногда встречаются такие явления, которые как будто специально созданы природой для проверки правильности теории. В отношении хромосомной теории примером может служить явление гтандроморфизма. Организмы, совмещающие в себе части тела разных полов — мужского и женского, называют гинандроморфами (гин— 9, андр— d" ). Гин-андроморфы существуют у тех видов, у которых четко выражен половой диморфизм (насекомые, птицы, человек), но встречаются они редко.
При латеральном гинандроморфизме, например у дрозофилы, одна половина тела имеет признаки женского пола, а другая — мужского (см. рис. на стр. 288). Как может возникнуть такой организм? Цитологические исследования показывают, что ткани гинандроморфа химерны: женская половина несет две Х-хромо-сомы, а мужская — одну.
На приведенном рисунке показан случай, когда у гинандроморфа рецессивный, сцепленный с полом ген white проявился на мужской стороне тела и не проявился на женской. Почему это так?
У гинандроморфа, возникшего из зиготы w+w, при первом делении дробления в силу каких-то необычных условий одна из Х-хромосом, несущая ген w+, в одной из дочерних клеток (бла-стомеров) утрачивается. Тогда две дочерние клетки окажутся
W
неодинаковыми в отношении Х-хромосом: одна ~. а вторая w.
Половина тела мухи, развившаяся из первой клетки, окажется женской и с красным глазом, а из второй разовьется половина тела с признаками мужского пола и с белым глазом,
поскольку рецессивный ген w, содержащийся в единственной ^-хромосоме, будет в гемизиготном состоянии.
Таким образом, и цитологический, и генетический анализ показывает, что в данном случае причиной гинандроморфизма может быть элиминация одной из ^-хромосом.
Кроме этого типа гинандроморфизма, который можно назвать монозиготным, известен также дизиготический гинандроморфизм. Он обнаружен у бабочек — Abraxas, тутового шелкопряда и у дрозофилы. Например, иногда в яйцеклетке тутового шелкопряда (самка гетерогаметна) образуются два женских пронуклеуса, один из которых кроме аутосом (обозначим их А) содержит Х-хромосому (Х+А), а другой — У+А. При полиспермии оба пронуклеуса будут оплодотворены разными спермиями, тогда в одном из бластомеров будет ХХ + АА, а в другом — ХУ+АА. Это и приведет к развитию дизиготного гинандроморфа. Аналогично может возникать гинандроморф у дрозофилы, только здесь различия между бластомерами получаются за счет разных сперматозоидов (самцы гетерога-метны).
Исключения из хромосомной теории определения пола. По мере накопления фактов хромосомная теория определения пола не только находила подтверждение, но и встречала некоторые трудности. Оставался открытым вопрос о том, не являются ли половые хромосомы индикаторами пола, вторично-половыми признаками?
Анализ исключительных особей у дрозофил, которые были получены в опытах Бриджеса, как результат нерасхождения половых хромосом (см. гл. 8) показал, что особи, имеющие, кроме аутосом, ХКУ-хромосомы (ХХУ+АА), являются самками, а особи ХО+АА — самцами. Эти факты убедительно говорили о том, что половые хромосомы отнюдь не индикаторы пола. Но как же они определяют пол, если особи ХУ+АА и ХО+АА являются самцами, а ХХ+АА и ХХУ+АА самками? Очевидно, дело обстоит не так просто, как это казалось вначале.
Дата добавления: 2020-01-07; просмотров: 284; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!