СТРОЕНИЕ ХРОМОСОМ И ИХ РЕПРОДУКЦИЯ
ГЕНЕТИКА
С ОСНОВАМИ СЕЛЕКЦИИ
|
|

М. Е. Лобашев
К В. Ватти
М. М. Тихомирова
ГЕНЕТИКА
С ОСНОВАМИ СЕЛЕКЦИИ
Лобашев М. Е. и др.
Л 68 Генетика с основами селекции. Учебник для пед. ин-тов. М., «Просвещение», 1970
431 с илл.+ l накидка на мел. бум.+ 1 цвет, накидка.
Перед загл. авт.: М. Е. Лобашев, К. В. Ватти, М. М. Тихомирова.
Учебник для пединститутов Утвержден Министерством просвещения РСФСР
20—69
ИЗДАТЕЛЬСТВО «ПРОСВЕЩЕНИЕ». МОСКВА 1970
|
|
| C$A |
| они |
| радиационная | | логическая генетика |
| химическаяШ ! химическая генетика / м \ генетика |
| логическая генетика |
| генетика микро организмов |
| частная генетика растений генетика . цитоплазмы . |
| генетика хромосом |
| генетика гамет |
| генетика [ соматических } клеток |
 генетика растений
генетика растений
[ эволюционнаяЫравнительная ^ — *-■ ц повед ения | Р
■ генетика У , генетика ^"\ * \ *\ ' е " с '" ка
| генетика животных |
| 1 1 |
космическая \_4 илогенегика ^ Ионтогенетика
Генетина
| фнзноло гнческая генетика |
Экологиче екая i генетика
| палеонто ( логическая генетика |
генетика простейших
Частная генетика животных
Генетика
Пола
| популяци онная генетика |
математи
ческая
| цитогенетика |
генетика
цитология J эмбриология
Карно систематика i
селекция
систематика
Предисловие
Биологическое образование становится насущной необходимостью каждого культурного человека. Это вызвано тем, что биология, наряду с физикой, химией и математикой, встала в ранг точных научных дисциплин и раскрывает новые горизонты в развитии производительных сил общества, в сельскохозяйственном и промышленном производстве.
Значение биологии как науки возросло благодаря тому, что одна из ее отраслей — генетика совершила настоящую революцию в познании и в методах изучения основных явлений природы, а именно воспроизведения клетки, наследственности и изменчивости. Основные загадки природы: возникновение жизни, развитие живой материи и использование органической энергии— могут быть разгаданы при условии познания трех указанных выше явлений. Для решения стоящих задач в настоящее время и концентрируются усилия биологов-генетиков, химиков, физиков, математиков и производится синтез знаний, добытых естествознанием в целом.
Учебник написан в объеме, соответствующем утвержденной программе для студентов педагогических институтов. Здесь даются лишь основные достижения современной генетики. Но генетика— быстро развивающаяся наука, и поэтому авторы стремились подчеркивать нерешенность тех или иных проблем.
Авторам хотелось бы предупредить читателей-студентов, что изучение генетики требует основательной общебиологической подготовки политологии, эмбриологии, биохимии, физиологии и систематике и знания других точных естественных наук (химии, физики и математики). Совершенно противопоказано отрывать рассмотрение генетических закономерностей от общебиологических.
Часто общебиологические и генетические закономерности, открытые на животных, противопоставляются таковым у человека. Человека от животных отличает вторая сигнальная система и социальные условия жизни, но биологические и генетические закономерности у них общие. Поэтому те генетические
7
закономерности, которые рассматриваются в книге на примере животных, растений и микроорганизмов, являются, как правило, универсальными и для человека. Это замечание мы считаем необходимым, поскольку педагог, зная огромные возможности социального воспитания и обучения детей, не должен забывать и об их генетическом разнообразии, что требует индивидуального подхода. Своевременное выявление генетической потенции ребенка может дать обществу гения, а больного — вылечить. В силу этих обстоятельств педагог не только дает генетические знания, но и сможет в свое время, когда будет разработано надежное тестирование, с большим успехом использовать их для выявления интеллектуального потенциала общества.
Авторы
Введение
Генетика изучает два основных свойства организмов — наследственность и изменчивость.
Наследственность — неотъемлемое свойство организмов передавать при размножении свои признаки и особенности развития потомству. Благодаря наследственности организмы некоторых видов оставались относительно неизменными в течение сотен миллионов лет, воспроизводя за это время большое количество поколений. Например, современный опоссум (Didelphys) мало чем отличается от опоссума раннего мелового периода, так же как кистеперая рыба латимерия (Latimeria) от своих девонских предков.
Организмы группируются в определенные систематические единицы: виды, роды, семейства и т. д. Эта системность возможна лишь при наличии наследственности, сохраняющей не только черты сходства внутри каждой группы организмов, но и различие между ними.
Обеспечение константного сохранения признаков в ряду последовательных поколений лишь одна из сторон наследственности; вторая сторона-—это обеспечение определенного типа развития и характера обмена веществ в онтогенезе. Каждому виду организмов свойственна определенная последовательность фаз и стадий развития. Так, например, дробление зиготы у человека начинается в яйцеводе, а на 5—6-й день после оплодотворения происходит имплантация, затем дифференцируются отдельные ткани, а потом уже закладываются органы. И все это происходит в соответствии с программой, которая записана в клетке, т. е. определяется наследственностью.
Мостиком, связывающим два поколения, т. е. материальной основой наследственности, являются: яйцеклетка и спермий при половом размножении и отдельная соматическая клетка при бесполом.
Клетки организмов не содержат готовых зародышей признаков взрослых особей: они несут в себе только задатки, возможности развития признаков и свойств, называемые генами. Ген — единица наследственности, определяющая отдельный элементарный признак, касается ли последний структуры белковой молекулы или элементарной реакции организма.
При изучении наследственности как одного из свойств живого следует различать два понятия: собственно наследственность и наследование. В понятие наследственности входит свойство генов детерминировать построение специфической белковой
9
молекулы, развитие признака и план строения организма. Наследование же отражает закономерности процесса передачи наследственных свойств организма от одного поколения к другому. При половом размножении наследование осуществляется через половые клетки, при бесполом и вегетативном — посредством деления соматической клетки.
В связи с этим и механизмы передачи могут быть различными.
Потомство, развивающееся в организме матери, может приобрести от нее в период беременности некоторые ненаследственные свойства (например, инфекционные болезни). Такие признаки получили название врожденных. У животных, обладающих нервной системой, мы встречаемся с особым типом функциональной преемственности приспособительных реакций между поколениями, когда потомство в порядке подражания родителям или в процессе воспитания вырабатывает те же условные рефлексы, которые приобрели родители в индивидуальной жизни. Поскольку в основе этой преемственности лежит механизм условного рефлекса, она может быть названа сигнальной наследственностью. Сигнальное наследование возникло в процессе эволюции как специальный механизм передачи индивидуального приспособления. Именно этот тип наследования лежит в основе процессов обучения и воспитания и обусловливает прогрессивное развитие человеческого общества.
Однако в генетической литературе довольно часто термин «наследственность» употребляется в широком смысле слова и включает как понятия наследственности и наследования в строгом смысле, так и другие формы преемственности между поколениями. В таком случае можно определить наследственность как свойство организмов обеспечивать материальную и функциональную преемственность между поколениями, а также обусловливать определенный характер индивидуального развития и план строения организма в соответствующих условиях внешней среды.
Наряду с явлением наследственности в предмет исследования генетики входит изучение изменчивости. Изменчивость является отражением нестабильного сохранения наследственных свойств организма. Она заключается в изменении генов и их комбинировании, а так же в изменении их проявления в процессе индивидуального развития организмов. Таким образом, наследственность сохраняет не только сходство, но и различия организмов в ряду поколений. Наследственность и изменчивость— два основных фактора, обеспечивающих эволюцию органических форм на Земле.
Современное изучение наследственности и изменчивости ведется на разных уровнях организации живой материи: молекулярном, хромосомном, клеточном, организменном и популяци-
10
онном. Это исследование осуществляется несколькими путями (методами), главным из которых является генетический анализ.
Систему скрещиваний в ряду поколений, дающую возможность анализировать закономерности наследственности и наследования отдельных свойств и признаков организмов при половом размножении, а также изменчивость генов и их комбинаторику, называют гибридологическим анализом. Это основной метод генетического анализа. Он включает в себя элементы математической статистики. Кроме того, в генетический анализ входит ряд других вспомогательных методов, заимствованных из эмбриологии, цитологии, физиологии и др.
Материальные основы наследственности изучают с помощью цитологического метода. Можно сказать, что этот метод служит для исследования «анатомии» наследственности. Изучение структуры клетки ведут с помощью световой и электронной микроскопии, рентгеноскопии и других приемов. Все шире для изучения материальных основ наследственности привлекаются цитохимические, биохимические, биофизические и физиологические методы. Сочетание гибридологического анализа с цитологическим составляет самостоятельный метод — цитогенетиче-ский.
Изучение действия гена и его проявления в индивидуальном развитии организма — один из разделов генетики, называемый феногенетикой, нам представляется правильнее называть этот раздел онтогенетикой. В онтогенетике применяются самые различные приемы анализа действия генов: трансплантация наследственно различных тканей,пересадка ядер из одной клетки в другую, методы культуры тканей, эмбриологический анализ развития, иммунологические реакции и т. д.
Таким образом, генетика изучает наследственность и наследственную изменчивость в трех основных аспектах: поведение генов в процессе размножения организмов, его материальную структуру, изменчивость и функцию (действие) гена в онтогенезе.
Официальной датой рождения генетики принято считать весну 1900 г., когда трое ученых, независимо друг от друга, в трех разных странах, на разных объектах, пришли к открытию некоторых важнейших закономерностей наследования признаков в потомстве гибридов. Г. де Фриз (в Голландии) на основании результатов работы с маком и другими растениями сообщил «о законе расщепления гибридов»; К. Корренс (в Германии) установил те же закономерности расщепления на кукурузе, а Э. Чермак (в Австрии) — на горохе.
Наука почти не знает неожиданных открытий. Это объясняется тем, что развитие ее обязано коллективному творчеству. Так случилось и с открытием законов наследственности. Оказалось, что трое ученых, открывших эти законы, всего-навсего
11
 «переоткрыли» закономерности, открытые еще в 1865 г. Грего-ром Менделем и изложенные им в статье «Опыты над растительными гибридами», опубликованной в «Записках общества естествоиспытателей» в г. Брно.
«переоткрыли» закономерности, открытые еще в 1865 г. Грего-ром Менделем и изложенные им в статье «Опыты над растительными гибридами», опубликованной в «Записках общества естествоиспытателей» в г. Брно.
Здесь нет необходимости излагать историю развития генетики, так как все содержание учебника посвящено этому вопросу, остановимся лишь на задачах и перспективах ее развития.
Генетика сейчас представлена большим количеством разделов, таких, как цитогенетика, генетика животных и человека, генетика микроорганизмов и простейших, генетика растений, математическая генетика, популяционная генетика и др.
Такая многосторонняя дифференциация генетики объясняется двумя обстоятельствами: во-первых, тем, что наследственность и изменчивость — основа возникновения и развития жизни на Земле; во-вторых, ее огромной важностью для развития сельскохозяйственного производства, медицины, а также для познания умственной и психической деятельности
человека.
Именно эти моменты определили прогресс генетики в очень короткий период истории естествознания.
Важным обстоятельством, подготовившим почву для рождения генетики и ее дифференциации, послужило быстрое развитие биологии как экспериментальной науки, в особенности систематики, эмбриологии, цитологии, селекции и др. Бурное развитие животноводства и племенного дела, растениеводства и семеноводства во второй половине XIX в. также породило повышенный интерес к явлениям наследственности и изменчивости.
Развитию науки о наследственности и изменчивости особенно способствовало учение Ч. Дарвина о происхождении видов, которое внесло в биологию исторический метод исследования эволюции организмов.
Современные задачи генетики вытекают из установленных общих закономерностей, характеризующих наследственность и изменчивость. К этим задачам относится изучение механизмов изменения гена, воспроизведения генов и хромосом в каждом клеточном делении, действия генов и контролирования ими элементарных реакций и образования сложных признаков и свойств в целом организме. Кроме того, из необходимости познания эволюции органической природы вытекает необходимость изучения взаимосвязи процессов наследственности, изменчивости и отбора.
Задачи современной генетики состоят не только в исследовании указанных теоретических проблем, раскрывающих перспективы и потенциал науки для познания кардинальных явлений природы. Перед генетикой стоят также и более близкие задачи, важные для достижения многих практических целей.
12
Сорт растения или порода животного — средство производства в сельском хозяйстве. Высокопродуктивные сорта растений и породы животных повышают производительность труда. Хотя выведение сортов и пород — задача самостоятельной науки — селекции, последняя не может развиваться без знания законов наследственности и изменчивости. Генетика раскрывает новые пути для селекции.
Генетика важна для решения многих медицинских вопросов. Так, по расчетам генетиков, на 3,5 млрд. человек, населяющих земной шар, 10 млн. человек в каждом поколении могут быть поражены различными наследственными болезнями. К их числу относят ряд тяжелых заболеваний нервной системы (эпилепсия, шизофрения), эндокринной системы (кретинизм), крови (гемофилия, некоторые анемии) и т. д. Ранняя диагностика наследственных болезней позволяет более успешно разрабатывать методы предупреждения их развития. С помощью новейших цитологических методов развертываются широкие исследования генетических причин различного рода заболеваний и их ранней диагностики, благодаря чему возник новый раздел медицины — медицинская цитогенетика.
В настоящее время можно назвать как вновь создающийся раздел генетики педагогическую генетику. Предметом ее изучения должна стать генетическая детерминация психологии и интеллектуальных способностей детей. Свои способности и психологические особенности дети наследуют от родителей так же, как любые другие признаки. Только проявление способностей, памяти и ассоциаций, в основе которых лежит физиологический механизм высшей нервной деятельности, описанный И. П. Павловым, обусловлено более сложным взаимодействием наследственности, социальной среды, воспитания и тренировки.
В разнообразии способностей детей учитель убеждается при первом же знакомстве с классом. Конечно, роль воспитания в проявлении наследственных способностей детей значительно большая, чем влияние внешней среды на формирование морфологических признаков. И тем не менее очевидно, что дети по своим способностям к отдельным видам деятельности неравноценны не только в разных семьях, но и внутри одной семьи. Педагогическая генетика, изучая наследственные способности детей, их возрастную изменчивость, корреляции различных способностей, роль сигнального наследования и т. д., должна давать рекомендации педагогике. Преподаватель иногда ориентируется на среднюю оценку способностей всех детей, и это приводит к тому, что в процессе обучения своевременно не выявляется одаренность ребенка к определенному виду деятельности, способности его не развиваются. Это имеет непосредственное отношение и к вопросу выбора будущей профессии оканчивающими школу.
13
Однако эта новая область не может развиваться без знания закономерностей общей генетики. Последняя необходима учителю и для понимания современного естествознания в целом.
Ионизирующие излучения, сопровождающие атомный взрыв, тредставляют двойную опасность для живых существ. При облучении поражаются не только соматические клетки (клетки тела), но и половые. Изменение первых ведет к различным заболеваниям облученного организма (лучевая болезнь), изменение вторых — к различным наследственным аномалиям у его потомства.
Развитие радиационной генетики в связи с исследованиями в космосе стало еще более необходимым. В космических полетах человек попадает под действие космических излучений. Отсюда возникает одна из проблем космической генетики — необходимость генетической оценки опасности этих излучений.
Особую роль генетика стала играть в фармацевтической промышленности в связи с открытием антибиотиков. Доступность антибиотиков (пенициллина, стрептомицина, биомицина и др.), спасших многие миллионы жизней, стала возможной благодаря успехам генетики: были использованы искусственно полученные наследственно измененные формы продуцентов антибиотиков, имеющие в сотни раз большую продуктивность.
С особой силой перед генетикой встает проблема производства аминокислот для кормления животных и питания человека. Решение этой задачи также возможно лишь на основе получения микроорганизмов с высокой продуктивностью аминокислот. Уже теперь в этом направлении получены определенные результаты: сотни тонн некоторых аминокислот (например, глютаминовой кислоты) получают ежегодно.
Современное естествознание раскрывает строение атомного ядра, но перед ним стоят еще более грандиозные задачи: овладеть энергией гена, определяющего и контролирующего синтез белковой молекулы, создать из неживой материи модель живой клетки и овладеть процессами воспроизведения клетки. Когда человечество подойдет к решению этих задач, его могуществу над природой не будет границ: откроются качественно новые возможности получения органического вещества и создания новых форм. Человек станет истинным конструктором жизни на Земле. В решении этих грандиозных задач естествознания генетика в комплексе с другими науками должна сыграть важную роль.
Как бы каждый из биологов ни сужал свои исследования, в конечном счете все они изучают законы эволюции животных, растений и микроорганизмов. Именно эта задача объединяет биологические дисциплины в единую систему — биологию. Роль генетики особенно велика, поскольку она изучает два основных фактора эволюции: наследственность и изменчивость.
14
Очевидно, в основе наследственности и изменчивости должны лежать сложные биохимические и физиологические процессы, без учета которых нельзя понять сущность наследственности и изменчивости. Так генетика вступает в тесный контакт с биохимией и физиологией.
Если индивидуальное развитие организмов определяется наследственными факторами — генами, то действие гена нельзя понять в отрыве от общих закономерностей онтогенеза. Так генетика вступает в связь с эмбриологией. Но было бы глубоким заблуждением полагать, что современная биохимия, физиология, эмбриология и другие биологические дисциплины могут решать свои задачи в отрыве от генетики.
Естествознание после столетнего периода дивергенции, расчленения на отдельные дисциплины, продолжая еще распадаться на более узкие специальности, вступило в эпоху объединения. Диалектика природы и методов ее познания привела к необходимости изучать не отдельные, изолированные процессы, а законы взаимосвязи явлений в природе.
Общий язык у биологии с такими точными науками, как физика, химия, математика, мог возникнуть лишь после того, как была установлена единица измерения биологического явления. Такой единицей измерения в биологии оказалась пока единица наследственности — ген. Благодаря этому именно генетика стала одной из наиболее точных и вместе с тем увлекательных дисциплин современного естествознания.
Большой вклад в развитие мировой науки внесла наша отечественная генетика. Крупнейшие советские генетики и цитологи открыли ряд важнейших закономерностей наследственности и изменчивости. Такие ученые, как И. Д. Чистяков, описавший деление клетки, С. Г. Навашин, открывший двойное оплодотворение у цветковых растений, Н. И. Вавилов, сформулировавший закон гомологических рядов наследственной изменчивости, И. П. Павлов, заложивший основы генетики поведения, И. В. Мичурин, создавший новые методы селекции плодовых растений, Г. Д. Карпеченко — автор метода преодоления бесплодия отдаленных гибридов, С. С. Четвериков — основатель популяционной генетики, Ю. А. Филипченко — зачинатель изучения частной генетики, А. С. Серебровский — пионер изучения тонкого строения гена, Н. К. Кольцов, развивший концепцию о химической природе гена, и заложивший основы селекции С. И. Жегалов, М. Ф. Иванов, А. П. Шехурдин, П. Н. Константинов, В. Я. Юрьев и ряд других, создали свои научные школы и разработали целые направления, которые заняли почетное место в истории мировой генетики. Ряд ныне живущих селекционеров (В. Н. Мамонтова, В. С. Пустовойт, П. П. Лукья-ненко) создали выдающиеся сорта растений — пшеницы, подсолнечника и других культур.

|

Раздел I. Материальные основы наследственности
Схема строения клетки по данным электронной микроскопии.
Каждый вид животных и растений сохраняет в ряду поколений характерные для него черты: курица выводит цыплят, свинья рождает поросят, рожь воспроизводит рожь и т. д., причем куда бы ни перевозили и в какие бы условия ни помещали представителей данного вида, если они сохранят способность размножаться, то воспроизведут себе подобных. Воспроизводство себе подобного потомства обеспечивает наследственность.
Что же является материальной основой наследственности, обеспечивающей эту преемственность?
Поскольку в основе размножения организмов лежит деление клетки, для ответа на поставленный вопрос необходимо рассмотреть строение клетки, ее химический состав, роль отдельных структур в функционировании и воспроизведении клеток.
Не менее важен вопрос и о том, каким способом наследственная информация передается от одного поколения к другому.
В природе существуют два способа размножения: бесполое и половое. Они принципиально отличаются друг от друга. При бесполом размножении одна клетка делится, воспроизводя целый организм. При половом размножении, как правило, две половые клетки (мужская и женская) соединяются и дают
Генетика с основами селекции
17
 Начало одной клетке (зиготе), которая затем уже делится и воспроизводит организм.
Начало одной клетке (зиготе), которая затем уже делится и воспроизводит организм.
Однако указанные два способа размножения имеют и общее: организм развивается из одной клетки.
Отличают еще вегетативное размножение, при котором новое поколение воспроизводится не из отдельной клетки, а из группы клеток эмбриональной или специализированной соматической ткани: отдельных органов (клубней, луковиц, корневищ) или из участков мицелия. Однако в основе вегетативного размножения, как и бесполого, лежит также процесс клеточного деления.
Глава 1. СТРОЕНИЕ КЛЕТКИ
Соматические и половые клетки многоклеточных животных и растений и одноклеточные организмы в принципе сходны по своему строению (см. рис. на стр. 16).
Они состоят из клеточной мембраны и протоплазмы. Протоплазма представлена ядром и цитоплазмой, содержащей различные органоиды (органеллы). Помимо общих для всех клеток структур, они обладают и рядом общих функциональных особенностей: использование и превращение энергии, синтез макромолекул из более простых веществ, самовоспроизведение и деление.
ЯДРО
Ядро является центром, управляющим жизнедеятельностью всей клетки и координирующим ее (см. главы 9, 15, 20). Оно имеет сложное строение, изменяющееся на разных фазах жизненного цикла клетки. В неделящейся клетке (интерфазе) ядро 'занимает приблизительно 10—20% ее объема. Оно окружено ядерной оболочкой (мембраной), пронизанной порами, через которые осуществляется обмен веществ между ядром и цитоплазмой.
Внутри ядра находятся хроматин, одно или несколько ядрышек и ядерный сок (кариолимфа, нуклеоплазма).
В ядерном соке в световом микроскопе можно различить сетчатую структуру с глыбками хроматина. По данным электронной микроскопии, эта сеть есть не что иное, как хромосомы, которые становятся хорошо различимыми только во время деления клетки.
Ядрышки — тельца, связанные с хромосомами, содержат большое количество рибонуклеиновой кислоты (РНК). Функция их еще недостаточно изучена. Имеются данные о том, что в них происходит синтез одной из РНК клетки, а именно рибосомной.
2. ЦИТОПЛАЗМА
Цитоплазма состоит из прозрачной жидкости гиалоплазмы и органоидов. Обязательным органоидом клетки является система мембран, служащих продолжением клеточной мембраны и имеющих гладкую или шероховатую поверхность из-за расположенных на них мельчайших гранул диаметром 100—120 А (рибо-
2*
19
 сомы). Эта система мембран получила название эндоплазмати-ческой сети. Она связывает клеточную и ядерную мембраны и имеет, как и они, липопротеиновую природу. По эндоплазмати-ческой сети — сети канальцев, образуемых мембранами, происходит движение веществ внутри клетки.
сомы). Эта система мембран получила название эндоплазмати-ческой сети. Она связывает клеточную и ядерную мембраны и имеет, как и они, липопротеиновую природу. По эндоплазмати-ческой сети — сети канальцев, образуемых мембранами, происходит движение веществ внутри клетки.
В цитоплазме находятся также и другие органоиды: митохондрии, рибосомы, аппарат Гольджи, лизосомы, центросома в клетках животных и низших растений, пластиды у растений и др.
Митохондрии бывают разной формы — палочковидные и гранулообразные. Для каждого вида организмов и типа тканей характерны различные по форме, величине и роли в биохимических процессах митохондрии. Их форма может изменяться в зависимости от функционального состояния клетки. Размеры митохондрий варьируют в значительных пределах: от 0,2 до 2—7 мк. Установлено, что они принимают участие в окислительном обмене клетки. Полагают, что в митохондриях происходят все реакции цикла Кребса. Митохондрии — поставщики энергии.
Рибосомы находятся в значительном количестве в цитоплазме (главным образом на поверхности мембран), а также в ядре. Они состоят из двух неравных по размеру частей. Общий размер их варьирует от 150 до 350 А, и в световом микроскопе они невидимы. Их особенностью является высокое содержание РНК (рибосомная) и белков; 80—90% всей клеточной РНК находится в рибосомах. Установлено, что рибосомы участвуют в синтезе клеточных белков под контролем ядра.
Аппарат Гольджи обеспечивает выделительную и секреторную функции клетки.
Лизосомы — тела, содержащие ряд ферментов и выполняющие функцию пищеварения внутри клетки: их ферменты способствуют расщеплению больших молекул на более мелкие составные части, которые могут окисляться ферментами митохондрий. Разрыв мембран, окружающих лизосомы, ведет к лизису содержащих их клеток.
Центросома (клеточный центр) состоит из двух компонентов: небольших телец — центриолей и центросферы — особым образом дифференцированного участка цитоплазмы. С центросомой связано формирование ахроматинового веретена, возникающего в период деления клеток. В клетках цветковых растений центросомы не найдены: здесь ахроматиновое веретено закладывается на полюсах деления в виде так называемых «полярных колпачков».
Пластиды (хлоропласты, лейкопласты, хромопласты и др.) характерны для цитоплазмы растительных клеток. Они осуществляют фотосинтез, синтез крахмала и пигментов, а также ли-пидов и пластидных нуклеиновых кислот.
Клеточная мембрана имеет сложное строение, которое приспособлено к выполнению определенных функций: защитная, избирательная проницаемость и активное втягивание частиц и молекул.
*
В целом можно сказать, что клетка является элементарной единицей жизни: в ней есть все необходимое для поддержания обмена веществ и размножения.
Краткое напоминание об общей структуре клетки было необходимо лишь для того, чтобы в последующем оценить роль отдельных ее элементов в наследственности.
Динамика структур в клеточном цикле и строение хромосом будут рассмотрены в следующей главе.
20
 Глава 2. ЦИТОЛОГИЧЕСКИЕ ОСНОВЫ БЕСПОЛОГО РАЗМНОЖЕНИЯ
Глава 2. ЦИТОЛОГИЧЕСКИЕ ОСНОВЫ БЕСПОЛОГО РАЗМНОЖЕНИЯ
В основе бесполого и вегетативного размножения организмов лежит универсальный процесс — деление клетки.
ДЕЛЕНИЕ КЛЕТКИ
Митоз. Деление клетки — центральный момент размножения организмов. В результате из одной клетки возникают две.
Этот процесс состоит из двух основных этапов: деление ядра— митоз (кариокинез) и деление цитоплазмы — цитокинез.
В жизненном цикле клетка проходит шесть последовательных стадий: интерфазу, профазу, прометафазу, метафазу, анафазу и телофазу (рис. 1,2). Все эти стадии составляют один митотиче-ский цикл, разделяемый на интерфазу и митоз.
Между двумя последовательными делениями клетки ядро находится в стадии интерфазы. Хотя интерфазу называют иначе фазой покоящегося ядра, на самом деле метаболические процессы в ядре в этот период, как будет видно дальше, совершаются с наибольшей активностью: клетка готовится к делению. В ядре в это время хорошо видна сетчатая структура, составленная из тонких нитей — хромосом.
В профазе — первой фазе митоза хромосомы спирализуются и становятся видимыми в световом микроскопе как двойные нити. Это убеждает нас в том, что в интерфазе осуществляется процесс удвоения, или репродукции, хромосом, при котором каждая из материнских хромосом строит себе подобную — дочернюю. Однако эти половинки, которые называются еще сестринскими хроматидами, в профазе не расходятся, а удерживаются вместе одним общим участком, называемым центромерой (кине-тохором).
В профазе хромосомы претерпевают процесс дальнейшей спи-рализации по оси, что приводит к их укорочению и утолщению. Важно подчеркнуть, что в профазе хромосомы располагаются по всему объему ядра.
В клетках животных в очень ранней интерфазе или даже еще в поздней телофазе предыдущего деления происходит удвоение центриоли, после чего в профазе начинается расхождение дочерних центриолей к полюсам клетки. Между центриолями появляется пучок тонких нитей веретена деления, совокупность которых называют ахроматиновым аппаратом.
22
 Нити веретена имеют белкопую природу, поэтому всегда перед их образованием в клетке идет интенсивный синтез и накопление белков.
Нити веретена имеют белкопую природу, поэтому всегда перед их образованием в клетке идет интенсивный синтез и накопление белков.
Существенными признаками окончания профазы являются исчезновение ядрышек и оболочки ядра, в результате чего хромосомы оказываются в общей массе цитоплазмы и нуклео-плазмы, которые теперь образуют миксоплазму.
Прометафаза характеризуется движением хромосом к экваториальной плоскости клетки. Это движение и распределение хромосом на экваторе веретена деления получило название метакинеза.
Метафазой называют стадию расположения хромосом в экваториальной плоскости, перпендикулярной оси веретена. Хромосомы, расположенные в этой плоскости, образуют экваториальную, или метафазную, пластинку. Каждая хромосома располагается таким образом, что ее центромера находится точно в экваториальной плоскости, а все остальное тело хромосомы может лежать и вне ее. При рассмотрении экваториальной пластинки с полюсов деления клетки хорошо видны все хромосомы, так что можно сосчитать их число и изучить форму. Нити веретена приобретают более плотную консистенцию, чем остальная масса цитоплазмы. Они прикрепляются к хромосомам таким образом, что к каждой центромере подходят нити от двух полюсов.
Анафазой называют следующую фазу митоза, в которой делятся центромеры, и сестринские хроматиды (которые теперь можно назвать уже хромосомами) расходятся к полюсам. При этом отталкиваются друг от друга в первую очередь центро-мерные участки, а затем расходятся к полюсам центромерами вперед и сами хромосомы.
Расхождение хромосом в анафазе начинается одновременно — «как по команде» — и завершается очень быстро. После расхождения хромосом их количество у каждого полюса оказывается одинаковым и точно соответствует общему числу хромосом исходной клетки. Благодаря такому способу деления ядра обеспечивается постоянство числа хромосом в клеточных поколениях.
В телофазе дочерние хромосомы деспирализуются и утрачивают видимую индивидуальность. Образуется оболочка ядра. Затем восстанавливается и ядрышко (или ядрышки), причем в том же числе, в котором они присутствовали и в родительских ядрах. Ядро реконструируется в обратном порядке по сравнению с теми изменениями, которые оно претерпевало в профазе.
Цитокинез. Пластиды размножаются путем деления и, по-видимому, не возникают в клетке заново. Предполагают, что и митохондрии размножаются путем деления. Вообще же о ме-
ханизмах репродукции органоидов клетки известно еще очень мало. Есть сведения, что процесс этот протекает асинхронно, а скорость деления частиц не совпадает со скоростью деления ядра. При цитокинезе распределение их по дочерним клеткам не подчиняется строгой закономерности, так как, по-видимому, не существует специального механизма, контролирующего этот процесс. В силу этого дочерние клетки могут получать неравное число одноименных органоидов. Случайное распределение органоидов между дочерними клетками не нарушает их нормальную жизнедеятельность. Очевидно, это происходит потому, что количество одноименных взаимозаменимых частиц в клетке очень велико.
Деление тела клетки — цитокинез начинается вслед за делением ядра. В животной клетке деление происходит путем перешнуровывания цитоплазмы по экватору материнской клетки от периферии к центру. В растительной клетке формирование клеточной перегородки идет при участии веретена за счет так называемого фрагмопласта от центра к периферии. Этим заканчивается митоз.
Продолжительность всего митотического цикла зависит от вида организма, типа ткани, физиологического состояния организма, внешних факторов (температуры, светового режима и др.) и колеблется в пределах от 30 минут до 3 часов. Скорость прохождения отдельных фаз митоза также изменчива.
Причины, определяющие готовность клетки к делению, и пусковой механизм до сих пор остаются невыясненными.
Кроме митоза известны и другие типы деления. Они встречаются, как правило, только в дифференцированных тканях. Рассмотрим некоторые из них: амитоз, эндомитоз, политению.
Амитоз. Прямое деление ядра, без образования ахромати-нового веретена, называют амитозом. Деление происходит путем перешнуровывания ядра на две части; иногда из одного ядра образуется сразу несколько ядер (фрагментация). Амитоз постоянно встречается в клетках ряда специализированных и патологических тканей, например: в крахмалообразующих клетках картофеля, в клетках мышц при регенерации, в раковых клетках, у простейших и др.
Эндомитоз. Эндомитозом называют такой процесс, при котором репродукция хромосом в клетке не сопровождается делением ядра. Вследствие этого в клетке происходит умножение числа хромосом, иногда в десятки раз по сравнению с исходным. Эндомитоз встречается в интенсивно функционирующих клетках различных тканей как растений, так и животных.
Политения. Иногда воспроизведение хромосом происходит без увеличения их числа в клетке. Кажднч хромосома многократно удваивается, но дочерние хромосомы остаются связанны-
24
ми
 ми между собой. Это явление называется политенией. Оно представляет собой частный случай эндомитоза. В результате политении диаметр хромосом заметно увеличивается. Число нитей в политенной хромосоме может достигать 1000—2000. В этом случае образуются так называемые гигантские хромосомы. Явление политении наблюдается в клетках ряда дифференцированных тканей и связано с особыми функциями ядра клетки. Поэтому политения характерна для тканей определенных органов, например слюнных желез двукрылых, и для клеток некоторых растений. Для того чтобы оценить значение происходящих при делении клетки процессов, необходимо более подробно рассмотреть строение и воспроизведение, т. е. репродукцию хромосом.
ми между собой. Это явление называется политенией. Оно представляет собой частный случай эндомитоза. В результате политении диаметр хромосом заметно увеличивается. Число нитей в политенной хромосоме может достигать 1000—2000. В этом случае образуются так называемые гигантские хромосомы. Явление политении наблюдается в клетках ряда дифференцированных тканей и связано с особыми функциями ядра клетки. Поэтому политения характерна для тканей определенных органов, например слюнных желез двукрылых, и для клеток некоторых растений. Для того чтобы оценить значение происходящих при делении клетки процессов, необходимо более подробно рассмотреть строение и воспроизведение, т. е. репродукцию хромосом.
СТРОЕНИЕ ХРОМОСОМ И ИХ РЕПРОДУКЦИЯ
Морфология и размеры хромосом. Общая морфология хромосом лучше всего выявляется на стадии метафазы или ранней анафазы, когда хромосомы наиболее укорочены и находятся в экваториальной плоскости. В это время хорошо видно, что они различаются по форме и величине (рис. 3). Форма каждой хромосомы определяется главным образом положением первичной перетяжки, где располагается центромера. Местоположение центромеры в разных хромосомах может быть различным, но оно постоянно и типично для каждой хромосомы.
Если центромера располагается в длинной хромосоме посередине, то в метафазе такая хромосома выглядит как равноплечая V-образная, или метацентрическая (рис. 3, 1, 6). Если центромера делит хромосому на два неравных участка, то образуются или слабо неравноплечая — суб мет ацентрическая (рис. 3, 2), или резко неравноплечая — акроцентрическая хромосома (рис. 3, 3, 4, 5). Предполагают, что центромера никогда не бывает на самом конце хромосомы. Концевые сегменты хромосом названы теломерами. Участок хромосомы, располагающийся ближе к центромере, называют проксимальным, а отдаленный ■— дистальным.
Кроме первичной перетяжки, хромосома может иметь вторичную перетяжку, не связанную с прикреплением нити веретена (рис. 3, 7). Эта перетяжка в хромосоме связана с формированием ядрышка и называется ядрышковым организатором. Выяснено,, что этот участок хромосомы имеет сложную структуру и ответствен за синтез рибосомной РНК. Иногда вторичная перетяжка может быть очень длинная, и тогда она отделяет от основного тела хромосомы небольшой участок, называемый спутником. Такие хромосомы называют спцтничными (рис. 3,5).
Как уже бь!лО сказано, К Центромере 6 метафазе прикрепляется нить веретена, разводящая хромосомы к полюсам. В случае разлома хромосомы бесцентромерный ее участок (ацентрический фрагмент) способен репродуцироваться, но восстановить центромеру не может. В силу отсутствия центромеры такой ацентрический фрагмент не имеет ориентации и при клеточном делении чаще всего утрачивается. Фрагмент сохранится лишь в случае, если он прикрепится к другой хромосоме, имеющей центромеру.
 Однако известны палочковидные хромосомы, к которым нити веретена могут прикрепляться по всей их длине. Такие
Однако известны палочковидные хромосомы, к которым нити веретена могут прикрепляться по всей их длине. Такие
| Типы метафазных хромосом: /, 6 — метацентрические (равноплечие); 2 — субметацентриче-ская (слабо неравноплечая); 3, 4, 5 — акроцентрические (резко неравноплечие); 7—акроцентрическая со вторичной перетяжкой; 8 — спутничная. Светлыми кружками обозначены центромеры. |
хромосомы имеют диффузную центро меру.В этом случае фрагменты разорвавшейся хромосомы могут нормально расходиться в анафазе. Природа этого явления остается пока малоисследованной.
Хромосомы различаются не только по морфологии, но и по величине. Длина их варьирует от 0,2 до 50 мк; диаметр от 0,2 до 5 мк (или 200—500 А). Однако длина каждой определенной хромосомы относительно постоянна. Таким образом, каждая хромосома индивидуальна.
Учитывая морфологию и величину хромосом, в клетке их можно точно идентифицировать, а для удобства изучения присваивать им определенные номера, что и было сделано для хромосом человека (см. гл. 29) и некоторых других организмов.
Структура хромосом. Структура хромосом делящихся клеток начинает вырисовываться в профазе. Как уже говорилось, в ранней профазе хромосомы имеют вид тонких двойных нитей {сестринские хроматиды). На стадии метафазы в световом микроскопе видно, что хромосомы состоят из 4 нитей, которые были названы полухроматидами. С помощью электронной микроскопии было показано, что полухроматида не
26
27
| Световой микросноп |
Электронный микроскоп
| Рентгено - структурншй анализ |
Полухро - матида ,-:
| Хромосома |
| Хроматида |
' ЭС*ЗАЛ^^иОДД~
| Диаметр одной нити Число молекул j 2 jfj ДНК на нить |
200-250 А I 00- I 25 A J 0- WA
8 4- 2
| Схема микроскопической, |
является предельной элементарной структурной единицей хромосомы. Была высказана гипотеза, что каждая хромосома состоит из многих нитей — хромонем. Диаметр тончайшей нити, по данным рентгеноструктурного ана-
о
лиза, может быть около 30 А. Многонитчатая структура хромосомы хорошо видна на рисунке 4. Согласно этой модели, общее число нитей в хромосоме — 64.
Однако высказана и другая гипотеза, согласно которой хромосома состоит только из одной нити, способной за счет спира-лизации и складывания создайать политенную структуру. Какая из этих гипотез справедлива, сейчас сказать трудно. Вопрос этот является предметом изучения цитологов.
Толщина хромосом в цикле митоза изменяется. Так, в профазе тонкие хромосомы начинают утолщаться, конденсироваться, и в метафазе они представлены в виде укороченных и утолщенных образований.
Каков же механизм сокращения и формирования хромосомы, характерной для метафазной пластинки? Было показано, что хромонемы в хромосоме по мере подготовки их к митозу претерпевают процесс спирализации. В ходе профазы спирали-зация распространяется по всей хромосоме, достигая максимума в метафазе. Поэтому в метафазе хромосомы выглядят очень компактными.
В настоящее время выяснено, что спирализация бывает двух порядков: мелкая и крупная, причем витки взаимно перпендикулярны. Оба типа спирализации идут почти одновременно (рис.5).
|
|

В телофазе наступает деспирализа-ция хромонем, и в интерфазе хромо-немы оказываются максимально раскрученными. Характер спирализации и деспирализации хромосом в мито-тическом цикле представляет закономерный процесс (цикл спирализации, рис. 6).
Следует обратить особое внимание на структурную дифференциацию хромосом по их длине.
| Схема строения метафазной хромосомы: / — морфология; 2 — внутренняя структура хроматиды, видимая при использовании специальных методов ослабления спирализа* ции. |
В профазе по всей длине хромосом заметны темноокрашивающиеся зернышки, или хромомеры. Положение их в каждой хромосоме постоянно, а в разных хромосомах—различно. Эти четкообразные утолщения (хромомеры) представляют собой не что иное, как спирализующиеся в первую очередь участки хромонем. Они оказываются наиболее уплотненными и поэтому в обычном световом микроскопе имеют вид темноокрашивающихся гранул.
При фиксации и окраске основными красителями разные участки (районы) дают разную реакцию. Одни участки интенсивно окрашиваются — их назвали гетерохроматиновыми, другие слабо окрашиваются—■ они названы эухроматиновыми.
Гетерохроматиновые участки разбросаны по всей длине хромосомы, но чаще располагаются вблизи центромеры и на концах, а также около ядрышка.
Существуют некоторые виды хромосом, которые преимущественно состоят из гетерохроматина. Гетерохроматиновые участки на протяжении всего жизненного цикла клетки, в том числе и в интерфазе, находятся в сильно спирализованном состоянии, а потому, очевидно, сильнее окрашиваются. Эухрома-тиновые участки в интерфазе деспирализуются, что может указывать на их более высокую метаболическую активность. По-видимому, глыбки хроматина в интерфазном ядре представляют собой не что иное, как гетерохроматиновые участки хромосом. При изменении окружающей среды они более лабильны, от них зависит частота разрывов и способность к восстановлению.
Дифференциация хромосом по длине особенно хорошо видна на гигантских хромосомах. Гигантские хромосомы в 100—200
28
29
 |
| 6. Схема спирализации хромо-нем в митотическом цикле: / — интерфаза, хромонемы слабо спиралнзованы (остаточные спирали); 2, 3, 4— профаза, усиление спирализации хромо-нем, образование двух хроматид; 5 — прометафаза, проявление четырех полухроматид; 6 — мета-фаза, максимальная спирализа-ция, выявляются как большая, так и малая спираль; 7 — анафаза; 8 — телофаза (одна из дочерних хромосом), деспирализа-ция хромонем. |
раз длиннее и содержат в 1000 раз больше хромонем, чем обычные мета-фазные хромосомы большинства соматических и половых клеток.
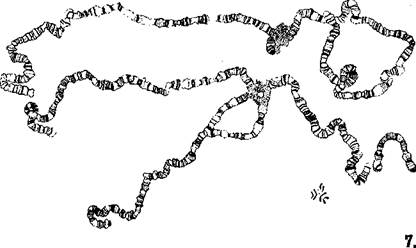
Впервые гигантские хромосомы были обнаружены Е. Бальбиани в 1881 г. в слюнных железах личинок мотыля (сем. Chironomidae). В дальнейшем оказалось, что такая структура хромосом характерна для ядер ряда соматических клеток личинок двукрылых — клеток кишечника, мальпигие-вых сосудов, слюнных желез, а также найдена у некоторых растений (в антиподах и синергидах) и у простейших.
Гигантские хромосомы возникают при эндомитозе. В этом случае 2 хромонемы после девяти последовательных удвоений образуют около 1000 нитей, плотно прилегающих друг к другу. Хромонемы гигантских хромосом постоянно находятся в частично и неравномерно деспирализованном состоянии, что обусловливает увеличение длины хромосом в 100—200 раз. Типичные гигантские хромосомы можно наблюдать в слюнных железах личинок дрозофилы (род Drosophila) (рис. 7). Строение и морфологические особенности этих хромосом видны в клетках при малом увеличении даже без специальной обработки на временных тотальных препаратах, но особенно хорошо — на окрашенных ацетокармином.
| Относительные размеры хромосом в ядрах клеток слюнных желез (гигантские) и в клетках ганглия (митотиче-ские) дрозофилы. |
К числу их особенностей относятся следующие. Если в любой соматической клетке хирономуса можно сосчитать 8 хромосом, то в клетке слюнной железы их только 4, так как одинаковые по морфологии и размеру хромосомы (одна отцовская, а другая материнская) обладают способностью объединяться, конъюги-ровать (соматическая конъюгация), что увеличивает еще больше толщину гигантских хромосом. Подробнее об этом будем говорить несколько позже.
Другая особенность состоит в том, что хромомеры многочисленных хромонем, плотно прилегая друг к другу, создают утолщения — диски, которые при окрашивании бывают более темными и создают впечатление поперечной исчерченности. Диски, так же как и хромомеры профазных хромосом,— более сильно спирализованные участки.
Размер и морфология дисков сильно варьируют, но для каждой хромосомы они постоянны и служат прекрасными маркерами при распознавании —идентификации хромосом. Между дисками хорошо видна политенность хромосомы.
Строение дисков изменяется в онтогенезе, что связано с функционированием хромосом, но об этом будет рассказано в главе 22.
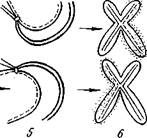
Другой моделью, на которой можно познакомиться с тонким строением хромосом и их функционированием, являются хромосомы типа «ламповых щеток». Вид этих хромосом действительно напоминает ершик, которым моют стеклянные пробирки, а в старину мыли стекла керосиновых ламп (рис. 8). Отдельные участки этих хромосом сильно вытянуты и образуют
30
31


 симметричные петли, перпендикулярные оси хромосомы (рис. 9). Такое состояние хромосом встречается в ооцитах рыб, амфибий, рептилий и птиц.
симметричные петли, перпендикулярные оси хромосомы (рис. 9). Такое состояние хромосом встречается в ооцитах рыб, амфибий, рептилий и птиц.
|
|

В отличие от гигантских хромосом «ламповые щетки» не являются политенными, а содержат сильно деспирализованные хро-монемы. Предполагают, что большая степень деспирализации связана с повышением метаболической активности хромосом в процессе роста ооцитов (см. главы 16, 21).
Тонкий электронномикроскопический анализ показал, что каждая хромонема по оси образует серию хромомер, из которых и выходят боковые петли — деспирализованные хромонемы, толщина их в самых тонких участках оказывается равной 100—
200 А.
Такова структура хромосом, как она представляется на современном уровне знаний.
| Схема строения отдельной петли, хромосомы типа «ламповых щеток». |
Химический состав хромосом. Изучение химического состава хромосом показало, что в основном они состоят из нуклеопро-теидов (90—92%)- Нуклеопротеид представлен дезоксирибонуклеиновой кислотой (ДНК) и белком — гистоном (или прота-мином). Кроме того, в хромосоме присутствует РНК, некоторое количество ионов кальция, магния, железа и др. и негистон-ные белки, иногда образующие комплекс
с РНК.
ДНК по своей природе является биологическим полимером, имеющим сложноор-ганизованную линейную структуру. Молекулярный вес ДНК очень велик, около 10 млн. и в отдельных случаях может доходить даже до 50—100 млн.
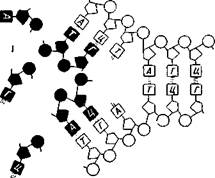
Молекула ДНК слагается из чередующихся мономерных единиц —дезоксирибонуклеотидов. В состав каждого нуклео-тида входит гетероциклическое азотистое основание (пуриновое или пиримидиновое), сахар —дезоксирибоза и остаток фосфорной кислоты (рис. 10). Универсально распространенными гетероциклическими основаниями, входящими в состав подавляющего большинства дезоксирибонуклеотидов, являются производные пурина — аденин и гуанин и производные пиримидина — цитозин и тимин,
| Ф |
| А |
|
|

| Строение отдельного нук-леотида (вверху) и фрагмента одиночной цепи ДНК (внизу): Ф — остаток фосфорной кислоты; Д— дезоксирибоза: П~~ пуриновое или пиримидиновое основание. |
Связь между нуклеотидами в цепи ДНК строго однотипна и осуществляется за счет образования диэфира фосфорной кислоты между определенными (3' и 5') гидроксилами соседних дезоксирибозных остатков. Таким образом, полимерная цепь ДНК состоит из последовательно чередующихся дезоксирибозных и фосфатных остатков. К каждому дезоксирибоз-ному остатку этой каркасной цепи присоединены боковые радикалы — пуриновое или пиримидиновое основание (рис. 11). В результате изучения химического состава ДНК у растений, животных и микроорганизмов было выяснено, что каждый вид характеризуется своим специфическим распределением пуриновых и пиримидиновых оснований, а также определенным молярным соотношением этих оснований. Отношение количества молей (гуанин + цитозин) к количеству молей (аденин+тимин) у разных видов колеблется в пределах от 0,45 до 2,8 (встречаются и более крайние отклонения). Оказалось, что последовательность нуклеотидов . в макромолекуле ДНК у разных видов неодинакова; это, как будет видно далее, имеет прямое отношение к наследственности.
Что же касается пространственной организации ДНК, то здесь обнаруживается поразительная однотипность. В одной молекуле ДНК объединяются две полинуклеотидные цепочки в виде двойной спирали с правым ходом винта (напоминающей винтовую лестницу), при этом пуриновые и пиримидиновые основания обеих цепей оказываются заключенными внутри пространства между витками спирали. Основания связаны друг с другом водородными связями. При этом пуриновому основанию одной цепи в норме соответствует пиримидиновое основание другой, и наоборот, а именно аденин всегда связан с ти-мином, а гуанин с цитозином. Таким образом, обе нити ДНК взаимно дополняют друг друга, что называется комплементар-ностью. Схема двуспиральной структуры ДНК представлена на рисунке 12.
В состав хромосом входит РНК, которая, как и ДНК, является полинуклеотидом. В ее состав входят 4 азотистых основания: аденин и цитозин, гуанин и урацил, Тимин в ней
32
33
° \ н н \
| н |
fv )"\ н ^ i
| Ф |
>- X \ \\ \\h-c-h I
- cNh h / ^ H /
A \ , C с—к
^ Ь Т 4 1
Ф \ н н К
ч с - н н н о
|
|
 а Да / Y *
а Да / Y *
ч н
О—Р=0 Н
\ l
Ф ХС—Н V=N
^ Ц н \
11.
Строение участка двойной иепи ДНК. Основания:
Д-аденин (пуриновое); Т - тимин (пиринидиновое); Г - гуаиин (пуриновое); Д - цито-
ззд (пДиримидиновое); Д - дезоксирибоза; Ф - остаток фосфорной кислоты.
| ПА |
 замещен урацилом, а дезоксирибоза— рибозой. В отличие от ДНК РНК имеет, как правило, однотяжевую структуру.
замещен урацилом, а дезоксирибоза— рибозой. В отличие от ДНК РНК имеет, как правило, однотяжевую структуру.
| 13 А |
Однако общая конструкция хромосомы на молекулярном уровне пока не ясна. Можно предполагать и то, что молекула ДНК в хромосоме непрерывна, и то, что хромосома состоит из многих коротких молекул ДНК, концы которых соединены связками, включающими негистонный белок и неорганические соли. Неясным остается и строение комплекса ДНК с ги-стоном.
В итоге можно сказать, что молекулярная, субмикроскопическая и микроскопическая структура хромосомы как целой системы находится сейчас в стадии интенсивного изучения.
| Схема двуспиральной структуры ДНК (модель Уотсо-на—Крика). |
Репродукция хромосом. Кардинальными вопросами в генетике, связанными с пониманием наследственности, являются: 1) в какой момент митоти-ческого цикла происходит воспроизведение хромосом и 2) каков молекулярный механизм этого воспроизведения.
Установлено, что важную роль в молекулярном механизме биосинтеза хромосом играет репликация ДНК как основного компонента хромосомы, хотя, конечно, знание механизма репликации ДНК не может полностью объяснить механизм удвоения хромосом.
Изучение синтеза ДНК в митотическом цикле показало, что у многоклеточных организмов он происходит в интерфазе, вот почему лишь условно можно называть интерфазу фазой покоящегося ядра (рис. 13).
Интерфазу делят на три периода, или фазы. В фазе, идущей вслед за прошедшим митозом, обозначаемой G \, ДНК не синтезируется, но осуществляется накопление продуктов (в том числе РНК и белка), необходимых для образования клеточных структур и следующего деления. Эта фаза названа пресинтети-ческой. Она самая длительная и лабильная. Продолжительность ее колеблется от 10 часов до нескольких суток.
Затем следует фаза синтеза ДНК (фаза 5), в течение которой количество ДНК в ядре клетки удваивается. Эта фаза длится 6—10 часов. В фазе S также осуществляется синтез РНК и белка.
35
Потом наступает постсинтетическая фаза (фаза G 2 ), когда ДНК не синтезируется, но идет синтез РНК и белков (в особенности ядерных) и накапливается энергия для следующего митоза. В клетке сохраняется удвоенное количество ДНК- Эта фаза длится 3—4 часа. Этим завершается интерфаза и наступает митоз, когда в результате деления клетки количество ДНК становится вновь равным исходному.
Успешному изучению продолжительности отдельных стадий интерфазы способствовало использование метода авторадиографии. Для этого в клетку вводили (в определенный момент) специфический предшественник ДНК — тимидин, меченный тритием (Н3). Он включается только в синтезируемую ДНК- Фиксируя клетки через разные промежутки времени после введения, устанавливали по появлению меченых ядер и изменению их процентного содержания продолжительность всех фаз цикла биосинтеза ДНК-
Каков же механизм удвоения ДНК? Предложено три схемы удвоения (редупликации) молекул ДНК (рис. 14): консервативный, полуконсервативный, дисперсионный.
При консервативном способе исходная двойная спираль ДНК остается неизменной и целостной в процессе синтеза и строит новую, себе подобную.
Полуконсервативный способ характеризуется тем, что цепи двойной спирали молекулы ДНК расходятся, не разрываясь, и каждая из одиночных цепей ДНК служит матрицей для образования комплементарной цепи. При дисперсионном способе в процессе удвоения молекулы ДНК составляющие ее цепи разрываются или разрушаются, так что после синтеза дочерних молекул последние включают в свой состав и случайным образом перекомбинированные фрагменты исходных молекул.
Полуконсервативное удвоение ДНК лучше других согласуется с моделью структуры ДНК, разработанной Дж. Уотсоном
|
|

и Ф. Криком, и соответствует экспериментальным данным генетики.
Согласно полуконсервативной схеме репликации ДНК сначала происходит разрыв водородных связей между пу-
13.
Митотический цикл. Интерфаза:
Qi — прееинтетическая фаза;
S — фаза синтеза ДНК; G 2 —
постсиитетическая фаза.

риновыми и пиримидиновыми осно- 14.
ваниями, образующими пары аденин- Схема различных способов
тимин и гуанин-цитозин. После раз- репликации (удвоения)
| ДНК: / — консервативный; 2 — полуконсервативный; 3 — дисперсионный; а—исходная молекула ДНК; 6—результаты одного цикла репликации; в— результаты второго цикла репликации. Черным цветом обозначены исходные молекулы ДНК. |
рыва двойная полинуклеотидная цепь раскручивается и каждая из образовавшихся одиночных цепей (моноспираль) строит около себя путем полимеризации комплементарную цепочку из мононуклеотидов, находящихся в кариоплазме. В результате образуются две молекулы ДНК, идентичные
исходной (рис. 15). Таким образом, в самой двойственности структуры ДНК, в комплементарное™ ее нуклеотидов, заключено важнейшее условие ее редупликации.
Принцип полуконсервативной схемы репликации ДНК, доказанный экспериментально, очевидно, приложим и для воспроизведения целых хромосом, которое, как уже говорилось, осуществляется в интерфазе. Это положение доказано опытами Дж. Тейлора, где методом авторадиографии была прослежена судьба исходных и дочерних хроматид в течение ряда последовательных митозов в клетках корешков конских бобов (Vicia faba) (рис. 16). При первом митозе, проходящем после включения метки, обе хроматиды каждой хромосомы оказались мечеными (рис. 16, 2). Однако во втором митозе, который шел при отсутствии в среде меченого тимидина, только одна из двух хроматид каждой материнской хромосомы содержала
3G
37
|
|

|
|

| Siiiii |

15. метку, поскольку включение метки во
| Схема, иллюстрирующая полуконсервативный механизм удвоения молекул ДНК: / — участок исходной молекулы ДНК; 2 — разрыв водородных связей между азотистыми основаниями двух тяжей; 3 —образование комплементарных цепочек из нуклеотидов окружающей среды (на рисунке — черные); 4 — две дочерние молекулы ДНК. Буквами обозначены азотистые основания: А — аденин, Г — ти-мин, Г — гуанин, Ц — цитозин. |
вновь редуплицированные хроматиды теперь уже происходить не могло (рис. 16, 5). Эти исследования позволили сделать вывод о том, что хромосома функционально состоит из 2-х единиц, после репродукции образуются 4 единицы, и разъединяются они таким образом, что каждая дочерняя хромосома содержит исходные и новые единицы. Иными словами, эти опыты доказали матричный полуконсервативый механизм воспроизведения хромосом. Эти данные были подтверждены исследованиями хромосом других растений из родов Bellevalia, Crepis, Allium, животных (хомячки) из сем. Cricetinae и человека, что говорит об универсальности этого механизма для высших форм.
Схема полуконсервативного удвоения хромосом хорошо согласуется со схемой полуконсервативной репликации молекулы ДНК, если допустить, что в момент, непосредственно предшествующий удвоению хромосомы, она состоит только из одной молекулы ДНК. Однако в настоящее время не ясно, как согла-
|
|
|
|

i
и
совать полуконсервативную схему уд- 16.
| Схема распределения исходных (немеченых—сплошная линия) и редуплицирован-ных Смеченых—пунктирная линия) хроматид и хромо-нем в митозе: /, 2, 3 — интерфаза, профаза, метафаза первого деления после введения в среду меченого ти-мидина; 4, 5, 6 — те же фазы второго деления при отсутствии в среде меченого тимидина. |
воения хромосомы с полуконсервативной репликацией молекулы ДНК в том случае, если исходная хромосома состоит из многих макромолекул ДНК. Предполагается, что репродукция хромосомы может осуществляться на молекулярном уровне, а не целой хромосомы, как это считалось раньше.
Метод авторадиографии показал также, что не все хромосомы в клетке редуплицируются в одно время, даже
отдельные участки каждой хромосомы редуплицируются в определенной последовательности (рис. 17). Такая асинхронность в редупликации различных частей хромосомы — еще одно из доказательств в пользу существования дифференциации хромосом по длине и, очевидно, она связана с их функционированием. Важным выводом из всего сказанного является то, что в процессе размножения хромосомы воспроизводятся таким способом, что образующиеся при делении клетки имеют их идентичные наборы.
3. ВИДОВАЯ СПЕЦИФИЧНОСТЬ КАРИОТИПА
Кариотип. Как было уже сказано, при изучении хромосом в экваториальной пластинке можно сосчитать их число в клетке, а анализ морфологии, структуры и размера хромосом позволяет их точно идентифицировать. Исследования хромосом клеток различных соматических тканей организмов одного вида показали, что каждому виду свойственно характерное число и состав хромосом. Набор хромосом соматической клетки, типичный для данной систематической группы животных или растений, называют кариотипом.
Каждому виду организмов свойствен определенный кариотип. Примеры кариотипов приведены на рисунке 18.
38
39

18. Если внимательно присмотреться к
Кариотипы разных видов кариотипу в любой соматической клет-
растений и животных, изо- ке, то можно заметить, что каждая
бражениые в одном масш- хромосома имеет парную, неотличи-
табе: мую по морфологии, структуре и раз-
со^ГТГа^пМаГОС2Ь-(мСу°ха Меру. Такая ПарНОСТЬ хрОМОСОМ ВОЗНИ-
So™^™Se,a(ncWr>cipifc кает за счет того- чт0 ПРИ образовании
ris); 4 -саранчовое (Gomphoce- ЗИГОТЫ В Процессе ОПЛОДОТВОреНИЯ ПО-(eUralislj?US): 5_ЖУК (GerdS "а" ЛОВИНу ХрОМОСОМ ПрИНОСИТ ЯЙЦеклетКЭ,
а половину — сперматозоид. Число хромосом в зрелых половых клетках называют гаплоидным и обозначают п. Соматические клетки — клетки тела многоклеточного организма содержат двойное число хромосом, его называют диплоидным и обозначают 2ге. Парные хромосомы, т.е. хромосомы, имеющие одинаковую морфологию, структуру и размеры, но имеющие разное происхождение (одна от матери, другая от отца) называют гомологичными-
На рисунке 19 изображен диплоидный набор хромосом скерды Crepis capillaris с указанием гомологичных пар.
Приведем примеры диплоидного числа хромосом у некоторых животных и растений.
Животные
Plasmodium malariae —малярийный плазмодий........................................... 2
Hydra vulgaris — гидра пресноводная ............................................. .... 32
Lumbricus terrestris —дождевой червь........................................................ 36
Bombyx mori —тутовый шелкопряд.................................................... 56
Pieris brassicae — капустная белянка...................................................... 30
Cyprinus carpio — сазан............................................................................ 104
Perca fluviatilis — окунь........................................................................... 28
Triturus vulgaris —тритон................................................................... • .24
Lacerta agilis — ящерица прыткая........................................................ 38
Columba livia —голубь.......................................................................... 80
Gallus gallus — курица домашняя....................................................... 78
Lepus cuniculus — кролик......................................................................... 44
Bos taurus — крупный рогатый скот................................................ 60
Anthropopithecus sp. — шимпанзе....................................................................... 48
Homo sapiens — человек......................................................... , . , , 46
40
|
|

19.
Диплоидный набор метафазных хромосом в клетке Crepis capillaris (2«=6). Одинаковыми цифрами помечены гомологичные хромосомы.
Растения
Abies, Picea, Pinus, Larix — пихта, ель, сосна, лиственница 24
Cucumis sativus —огурец.................................................... ......... 14
Ribes nibrum —красная смородина ........................... ......... 16
Malus silvestris —яблоня................................................... 34,51
Quercus robur —дуб обыкновенный............................. 24
Solanum tuberosum — картофель............................................ 48
| 8) В 7 В |
Secale cereale —рожь........................................................ 14+(0 —
Zea mays — кукуруза............................................... 20 -f (1 —
Количество хромосом в кариотипе не связано с уровнем организации животных и растений: примитивные формы могут иметь большее число хромосом, чем высокоорганизованные, и наоборот. Однако число и морфология хромосом в отдельных случаях могут служить показателем филогенетического родства видов. На этом принципе строится кариосистематика.
Хотя мы и говорим о законе постоянства числа и формы хромосом в наборе клетки для каждого вида организмов, однако следует отметить, что это постоянство относительно.
Клетки разных тканей даже одного организма в зависимости от выполняемой функции могут содержать разное число хромосом. Так, например, в клетках печени животных бывает большее, чем два, число наборов хромосом (т. е. An , 8 n ).
Некоторые виды растений могут быть представлены формами, отличающимися числом хромосом, кратным гаплоидному. Так, рожь ( Secale cereale) может иметь 2 п—14 хромосом или 28 хромосом (4 п). При этом все видовые признаки у обеих форм ржи сохраняются.
Добавочные хромосомы. Установлено, что у некоторых видов растений (кукуруза, рожь и др.), а также у животных, например у пресноводных тубеллярий и некоторых насекомых, имеются так называемые добавочные к диплоидному набору хромосомы.
В отличие от хромосом нормального диплоидного набора, названных хромосомами типа А, дополнительные хромосомы были названы типом В. В-хромосомы, в отличие от основных Л-хро-
41
мосом, более интенсивно окрашиваются, потому что, как правило, имеют больше гетерохроматина. В анафазе мейоза часто не наблюдается их равного распределения, поэтому в дочерние клетки может попадать неравное их число, с чем связана изменчивость кариотипа по числу Б-хромосом. Так, у кукурузы (Zea mays) количество их в клетке может варьировать от
1 до 34.
Наличие небольшого числа Б-хромосом заметно не сказывается на росте и морфологии растения, но накопление их в большом числе (более 10) вызывает депрессию роста, снижение плодовитости и различные аномалии в свойствах н признаках.
Указанные случаи отклонения числа хромосом от нормального диплоидного набора не дают основания отрицать правило постоянства числа и формы хромосом для каждого вида животных и растений.
* *
*
Итак, в результате митоза из одной клетки образуются две
дочерние.
Как было показано, основные компоненты клетки — хромосомы в ходе митоза репродуцируются таким образом, что обе сестринские хроматиды оказываются идентичными. Ахроматино-вое веретено обеспечивает точное распределение их в дочерние клетки, так что последние содержат такие же наборы хромосом, как и исходная клетка.
Органоиды распределяются между образующимися клетками случайно, но более или менее равномерно. Отсутствие механизма точного распределения их компенсируется, очевидно, их большим числом и полной взаимной заменяемостью.
Таким образом, цитологической основой бесполого размножения является митоз, в результате которого из одной клетки воспроизводятся две идентичные.
Дата добавления: 2020-01-07; просмотров: 786; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!