Сравнение в химическом отношении различных частей листьев липы и желатина 2 страница
Баруха-Рейд (1956) считает результаты Иеркса, Хека и Робинзона достоверными. Его интересовало явление так называемого латентного обучения. Одна особь дождевого червя ( Lumbricus terrestris ) пребывала в течение 20 час. в Т-образном лабиринте. В это время никакие реакции животного не подкрепляли. Затем исследовали, спустя какое время после начала применения удара электрическим током червь овладеет лабиринтом по сравнению с контрольными особями, к которым удар током применяли с самого начала пребывания их в лабиринте. Наблюдение велось всего на 6 особях (3 подопытных и 3 контрольных). На конце левого прохода лабиринта (запретное направление) пол был выстлан грубой наждачной бумагой. На правом конце находился сосуд с сырой землей и мхом. В конце левого прохода черви получали удар током. Каждый червь совершал 6 пробегов в день. Показатель решения задачи принят условно:
7 правильных реакций из 10. В первой группе (контрольной) условный показатель был достигнут после 45, 37 и 47 пробегов, в по-
50 Ян Дембовский
допытной группе — соответственно после 21, 22 и 23 пробегов. Разница была статистически достоверна.
В заключение я опишу "сутки дождевого червя". Вырыв норку, животное в продолжение всего дня сидит в ней неподвижно головой ко входу, довольно плотно закрытому листьями или прутиками. В сумерки червь оживляется и приступает к работе. Он выползает из .норки, вытягивая тело во всю длину (максимальное сокращение кольцевых мышц) и цепляясь задним концом за край норки. Передний конец тела совершает круговые движения, несколько приподнимаясь над землей и ощупывая все вокруг. Время от времени тело животного вытягивается в другом направлении и головной конец исследует новые районы. Натолкнувшись на какой-либо предмет, дождевой червь присасывается к нему. Вокруг ротового отверстия находятся мягкие губы, плотно прижимающиеся к данному предмету. Одновременно глотка, выдвинутая вперед в момент схватывания, отодвигается назад, действуя как поршень, создающий отрицательное давление. В результате тубы крепко присасываются к поверхности предмета. Пятясь обратно в норку, червь тащит предмет за собой. Маленький камешек, попадающий в норку, червь вдавливает в стенку, а большой камешек, подтащив к отверстию, бросает. То же самое делает он и с листьями: если после нескольких попыток ему не удается втянуть лист, то червь оставляет его на месте. Таким образом, в течение ночи над норкой скапливаются листья и другие предметы, закрывающие отверстие.
|
|
|
Ночью в состоянии максимального растяжения дождевой червь особенно чувствителен ко всяким воздействиям среды, особенно к прикосновениям, толчкам, химическим влияниям и свету. Поэтому наблюдать за червем можно, только соблюдая большую осторожность. Нужно очень медленно двигаться и освещать червей только слабым красным светом. Очень слабый белый свет притягивает дождевых червей. Иногда в лунную ночь наблюдаются далекие путешествия дождевых червей. Однако они неспособны отыскать брошенную ими норку и поэтому с наступлением дня роют новую. Утром черви прячутся в своих норках.
|
|
|
Это животное обладает очень разнообразными органами Чувств и на некоторые раздражители реагирует уже при минимальной интенсивности. Дождевой червь имеет различные средства для- познавания присущим ему способом окружающего мира; он может анализировать-влияния внешней среды и накапливать приобретенный опыт. Жизнь червя ка
жется нам монотонной и бедной только потому, что мы еще не изучили ее как следует. Очень сложное строение нервной системы, обилие и разнородность связей между ее элементами, изученных лишь в небольшой степени, показывают, что мы имеем дело с животным, которому присущи многообразные восприятия.
|
|
|
Ж.И. Резникова
ПОВЕДЕНИЕ МУРАВЬЕВ НА КОРМОВОМ УЧАСТКЕ*
ся их "язык". До сих пор неясно, могут ли муравьи использовать дистанционное наведение на источник пищи, подобно пчелам, какое количество информации они могут передавать и какие факторы определяют выбор того или иного способа обмена сведениями.
В данной главе приводятся лишь первые подходы к решению этих вопросов, отнюдь не претендующие на полноту исследования.
Функционирование такой сложной системы, как многовидовое сообщество муравьев, во многом определяется характером поведения и взаимодействия особей на кормовом участке. Появляется все больше данных в пользу того, что действия муравьев, главным образом, социально обусловлены (Захаров, 1972,1974, 1975). К настоящему времени многое известно о различных формах координации деятельности внегнездовых рабочих, а также о способах добывания пищи и об особенностях ориентации. Однако за пределами исследований остались некоторые важные вопросы поведения муравьев, познание которых необходимо для разработки концепции сообщества социальных насекомых.
Одна из наименее изученных сторон жизни муравьев — индивидуальное поведение особей и роль индивидуума в жизни семьи. До сих пор основное внимание исследователей привлекало групповое поведение. Оценка внутривидовой и межвидовой изменчивости индивидуального поведения муравьев пока практически отсутствует.
|
|
|
Нет экспериментальных работ, посвященных исследовательскому поведению муравьев в естественных условиях; в то же время это — важная форма деятельности, присущая всем животным. Многие этологи склонны рассматривать его как фундамент всего поведения (Шовен, 1972). Изучение исследовательского поведения в сравнительно-видовом аспекте может пролить свет на возможности использования биологического потенциала на уровне многовидовых сообществ муравьев. То же касается способности муравьев к научению, поскольку наши представления о сложности задач, с которыми в природных ситуациях сталкиваются муравьи, и о способах их решения еще явно недостаточны.
Наконец, едва ли не самой интригующей стороной социальной жизни муравьев являет-
* Резникова Ж.И. Межвидовые отношения муравьев. Новосибирск: Наука, 1983. С. 51—91 (с сокр.).
ИНДИВИДУАЛЬНОЕ ПОВЕДЕНИЕ МУРАВЬЕВ
Среди немногих работ, посвященных изучению индивидуального поведения муравьев, большинство проведено в лабораторных условиях и посвящено главным образом функциональному разделению особей в семье и различиям в уровнях их активности. Я. Добжанская (Dobrzanska, 1958), используя индивидуальное мечение, впервые поставила ряд экспериментов по выявлению связи поведения муравьев F . polyctena и F . sanguined с выполняемыми ими функциями. Оказалось, что наименее раздражимые особи несут функции, не требующие подвижности; другие выполняют работы, связанные с частой сменой задач и с постоянным активным движением.
Г. Веррон (Verron, 1974, 1976) исследовал локомоторную активность муравьев Lasius niger , Lasius flavus и Formica fusca , помещая их на разграфленный манеж и учитывая число делений, пробегаемых каждой особью, а также время, затраченное на передвижение и остановки. Оказалось, что часть особей отличается большой вариабельностью этих этологических признаков, остальных можно условно разделить на сильноактивных (в семье их меньшинство), средне- и низкоактивных. Сходные данные были получены на Tapinoina erraticum в результате наблюдений за транспортировкой куколок из чрезмерно нагретой в прохладную зону (Meudes, 1973).
У Myrmica laevinodis исследовались индивидуальные различия в уровне активности и агрессивности (Le Roux A. M., Le Roux С., 1979). Оказалось, что в среднем 50% рабочих совершенно неактивны и не выходят из гнезда, 30% активны, но не агрессивны, а 20% и активны и агрессивны, причем последняя группа может быть еще разделена на малоагрессивных и агрессивных.
В естественных условиях индивидуальное поведение муравьев изучалось в плане функциональной дифференциации особей (Oekland,
52 Ж . И . Резникова
1931; Kill, 1934; Stager, 1937; Otto, 1962;
Boulig, Janzen, 1969). Так, К. Хорстманн (Horstmann, 1973) выделяет среди внегнез-довых рабочих F . polyctena три профессиональные группы: восходители на деревья, специализированные охотники на почве и сборщики строительного материала. Собственно индивидуальной изменчивости поведения внегнездо-вых рабочих на кормовом участке посвящена только работа Ф. Бернарда (Bernard, 1971), описавшего различия в технике сбора семян у Messor barbara : большинство муравьев таскают семена одно за другим, пробираясь в густых зарослях, 8—9% срезают и тащат в гнездо целый стебелек с 20—60 семенами, и лишь 1—2% сборщиков умеют сбрасывать зерна вниз с вершины холма.
Соотношение различных элементов поведения у муравьев разных функциональных групп до сих пор остается неисследованным у подавляющего большинства видов. В частности, лишь в немногих работах применялся метод построения этограмм (Harkness M.L., Hark-ness R.D., 1976; Higashi, 1978; Brandao, 1978).
<...> Мы оцениваем индивидуальное поведение муравьев, согласно принципам фиксирования и классификации элементов поведения Е.Н.Панова (1978) по соотношению элементов поведения и, кроме того, по характеру траекторий передвижения на кормовом участке (Резникова, 1974 а, 1977 а; Резникова, Шиллерова, 1978, 1979). Большой вклад в эти исследования внесла О.А. Шиллерова (Богатырева) (1980). Вопросам индивидуального поведения муравьев разных видов посвящена ее дипломная работа (1980).
Наблюдения проводились в 1971,1976—1979 гг. в ассоциациях I и II*. Предварительно большую часть внегнездовых рабочих в выбранных семьях метили индивидуальными метками с помощью цветных нитролаков, смешанных с нитроклеем. Метка держалась на муравьях до полутора месяцев. Код составлялся так, чтобы стирание краски можно было контролировать:
например, желтая точка на груди муравья означала, что белой точке на левой стороне брюшка должна соответствовать красная точка на правой. Все кодовые сочетания заносились в журнал, и одновременно с мечением новых муравьев производилось подновление старых меток. Для индивидуального отлова муравьев проворных видов применялись конические колбочки с прорезанным в дне отверстием.
* Ассоциация — здесь: группа видов муравьев, обитающих на общей территории (прим. сост.).
Наблюдения проводили в периоды высокой активности муравьев с момента выхода из гнезда до возвращения в него. Для статистической обработки отбирали "полные" рейсы, исключая муравьев, потерянных наблюдателями до возвращения в гнездо. Два наблюдателя хронометрировали продолжительность поведенческих реакций муравьев с помощью двухстрелочного секундомера и одновременно вычерчивали траекторию передвижения особи на плане кормового участка. Для этого кормовой участок предварительно был разбит на квадраты со стороной 0,5м. Всего проанализировано 640 этограмм и траекторий муравьев, что соответствует примерно 100 ч наблюдений.
Мы старались проследить как можно большее число рейсов одной и той же особи за любой срок — от одного дня до месяца. Такие наблюдения в природе, насколько нам известно, не проводились. В то же время без них очень трудно представить себе место муравья в семье и изменчивость его поведения. Это оказалось сложной задачей: многие меченые муравьи теряли метку или терялись сами, погибали на кормовом участке, становясь добычей пауков или других муравьев, а чаще всего на неопределенный срок исчезали в гнезде. Поэтому мы очень дорожили теми особями, которые исправно выходили на кормовой участок и давали нам возможность судить о степени постоянства функций муравьев на кормовом участке и об их территориальном консерватизме. Таким муравьям мы давали имена и старались использовать каждую встречу с ними в своих целях. Иногда удавалось наблюдать до 20 рейсов одного фуражира в течение месяца.
Пользуясь классификацией Е.Н. Панова (1978), мы выделяли в качестве регистрационных элементов единицы поведения II уровня — несложные блоки локомоций (бег, транспортировка добычи, обследование травостоя) и "позы" (резкий поворот на месте, ощупывание предмета). Правда, в некоторых случаях трудно решить, к какому уровню принадлежит рассматриваемая единица поведения: ко II или к III. Так, обследование небольших площадок, травостоя и т. п. представляет скорее "комплекты поз", однако ввиду их несложности нам кажется нецелесообразным рассматривать их отдельно. Более или менее упорядоченные последовательности (ориентировочные окологнездовые, охотничьи рейды и т. п.) мы рассматривали в качестве поведенческих ансамблей.
Поведение муравьев на кормовом участке 53
Для количественной оценки траекторий (Резникова, 1980) были выбраны следующие параметры:
1) отдаление — расстояние от гнезда до наиболее удаленной от него точки траектории;
2) степень извилистости — отношение отдаления к общей длине пробега;
3) отношение числа самопересечений траектории к общей длине пробега;
4) максимальная ширина охвата участка траекторией;
5) площадь охвата участка траекторией;
6) амплитуда отклонения траектории от прямой, проведенной от гнезда до наиболее удаленной от него точки (среднее из 10 измерений, произведенных на равном расстоянии).
ВНУТРИВИДОВАЯ ИЗМЕНЧИВОСТЬ ИНДИВИДУАЛЬНОГО ПОВЕДЕНИЯ
Наблюдения за тремя семьями Formica cunicularia (Резникова, Шиллерова, 1978) позволили выделить две относительно автономных группы: трофобионтов и активных фуражиров* (соотношение их численности примерно 1 : 2). И те и другие могут заниматься транспортировкой строительного материала. Серии дальних рейсов (их бывает 3—5 за день) обычно начинаются и заканчиваются кратковременными выходами в пригнездовую зону.
Трофобионты, посещающие наземные колонии тлей на переферии кормового участка, используют в большинстве рейсов один и тот же путь. Около 90% времени они затрачивают на бег по поверхности почвы или по стеблям трав, остальное время приходится на чистку тела или ориентировочные реакции. Муравьи, посещающие кормовые пещерки (с подземными колониями тлей и цикадок) в средней зоне кормового участка, обладают более изменчивым и разнообразным поведением. Они тратят значительно больше времени (до 25%) на обследование территории (ощупывание мелких предметов, прикорневых частей растений, обследование почвы), что, возможно, связано с поиском новых кормовых пещерок. Эти муравьи изредка могут совершать охотничьи рейсы.
' Трофобионты — группа рабочих мурвьее с узкой специализацией —собиратели выделений тлей.
Активные фуражиры — рабочие муравьи, активно разыскивающие корм и производящие мобилизацию пассивных фуражиров к источникам корма (прим. сост.).
Активные фуражиры ("охотники") обрабатывают широкие секторы кормового участка и, видимо, хорошо знакомы со всей территорией в целом. На пути фуражира, несущего добычу, мы последовательно ставили преграды, перегоняя его таким образом в самый дальний сектор по отношению к тому, где этот муравей был впервые замечен. Из 14 особей лишь 2 затратили на возвращение в гнездо более 10 мин, остальные находили его без всяких затруднений. Ранее такие опыты, позволяющие предположить наличие "внутреннего плана" территории у активных фуражиров, проводились нами у Cataglyphis aemscens и А. Буркхалтером (Burkhalter, 1972), М. и Р. Хар-кнессами (Harkness М., Harkness R., 1976) у Calaglyphis bicolor .
Характер поведения и конфигурация траекторий наиболее лабильны у активных фуражиров. Траектории их отличаются большой извилистостью (0,11±0,07), тогда как при сборе пади** этот показатель близок к единице. Во время охоты наблюдаются и наибольшая площадь охвата территории — 9,62 ± 2,3 м2 (при сборе пади — 1,19 + 0,6 м2, при окологнездовом патрулировании — 1,6 ± 0,1) и максимальная амплитуда отклонения (соответственно 2,13 ± 0,2 м; 0,7 ± 0,3 м и 0,4 ± 0,08 м).
При поиске добычи муравьи либо передвигаются широким челноком, либо последовательно обследуют небольшие участки на разном удалении от гнезда. Поведенческий репертуар охотника включает обыскивание травостоя, ощупывание предметов, охоту и транспортировку добычи. Чаще всего муравей находит мелких насекомых (цикадок, клопов) в куртинах злаков или в укрытиях на поверхности почвы, сжимает их в челюстях и сразу транспортирует в гнездо. В поисках добычи активные фуражиры могут в течение нескольких дней или нескольких периодов активности обрабатывать один и тот же участок и, наоборот, в один день обследовать разные секторы территории.
Спектры поведенческих реакций муравьев во время охотничьих рейсов весьма изменчивы. Однако можно выделить и присущие каждому муравью особенности. Так, на рис. 1, а, б видно, что муравей Ml очень подвижен и почти не тратит время на чистку тела и обследование травостоя и мелких участков на поверхности почвы; напротив, у муравья М2не менее 30% времени уходит на обследование травостоя или ориентировочные реакции.
** Падь — сладкие выделения тлей, которыми питаются муравьи (прим. сост.).
54 Ж. И. Резникова
| а |
| б |
|
|

|
|
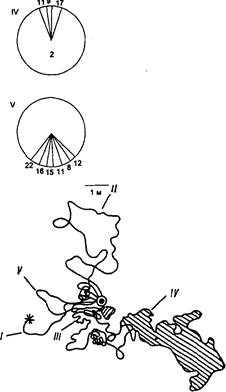
Рис. 1. Индивидуальные траектории и этограммы двух особей Formica cunicularia M,(a) и Мд (б) в разные дни и часы и усредненная этограмма для 50 активных фуражиров лугового муравья (в):
Элементы поведения для рис. 20—22: 1 — бежит по прямой, 2 — перемещается плавными зигзагами, 3 — бежит с кратковременными остановками, 4 — резко поворачивает, 5 -останавливается, приподнимаясь на передних ногах, б — подробно обследует небольшие участки почвы, 7 — стоит неподвижно, 8 — ощупывает предметы антеннами, 9 — стоит и шевелит антеннами, 10 — оглядывается, 11 — взбирается на возвышения и оглядывается, 12 — обследует травостой, 13 — перебирается через травинки, 14 — находит добычу, /5 -умерщвляет добычу, 16—транспортирует добычу, 17— соприкасается антеннами с другими муравьями, 18 — избегает контакта с другими муравьями, 19 — обменивается пищей (трофаллаксис), 20 — стычка с другими муравьями, 21 —чистит антенны, 22 — чистит тело и ноги; 1—V—номера траекторий движения и соответствующие им этограммы: для М1 16/ VIII в 17.05 (I), 16.35 (II) и 10.03 (III); для М2 24/VIII в 16.05 (I), 10.20 (II), 15/VIII в 17,02 (III), 26,VIII в 11.15 (IV) и 26/VIII в 8.15 (V); на траекториях заштрихованы области, которые муравьи тщательно обследовали
Поведение муравьев на кормовом участке 55
У F . pratensis поведение особей связано с глубокой специализацией. Для этого вида характерно четкое вторичное деление территории и наличие узкопрофессиональных групп фуражиров. Поэтому индивидуальная изменчивость поведения отдельных особей выражена значительно слабее, чем у F . cunicularia . При сравнении активных фуражиров оказалось, что в то время как в поведении каждой особи F. cunicularia содержится почти полный набор актов, выделенных нами у обоих видов, у F . pratensis в среднем их 6—8. Отличительная особенность поведения лугового муравья — большая доля времени, затрачиваемого на контакты с другими муравьями на кормовом участке (рис. 1, б).
Дата добавления: 2019-07-15; просмотров: 154; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!