Сравнение в химическом отношении различных частей листьев липы и желатина 3 страница
Formica uralensis экологически во многом сходен с F . pratensis (Stebaev, Rcznikova, 1972, Резникова, Самошилова, 1981), но организация кормового участка и поведение особей черноголового муравья совершенно иные (см. ранее). Как уже отмечалось, вторичное деление территории выражено очень слабо. Большинство внегнездо-вых рабочих обрабатывают разные зоны кормового участка и, по-видимому, могут осуществлять весь набор функций. Из 20 особей, наблюдавшихся многократно, лишь один муравей совершал только охотничьи рейсы, а остальные совмещали их с посещением кормовых пещерок и колоний тлей.
Типичный случай представлен на рис. 2. Муравей В 24/VII в 9.05 вышел для окологнездового ориентирования (траектория I). В 9.20 он выходил с охотничьим рейсом (траектория II), причем часто контактировал с другими особями, обследовал растительность, останавливался для ориентировки. В 18.53 он охотился несколько дальше от гнезда (траектория III). В другие дни этот муравей посещал кормовую пещерку на дороге (траектория IV). 20/VII он дважды выходил на охоту в разные зоны кормового участка (траектория V, VI), а также посещал кормовую пещерку (траектория VII). Помимо этого, он участвовал в столкновении с семьей F . picea , находящейся в 9 м от гнезда по ходу дороги. Характерно, что чем дальше муравьи отходят от гнезда, тем больший процент времени в их поведенческом репертуаре занимают ориентировочные реакции и меньший — контакт с другими муравьями. При повторных выходах на тот же маршрут доля ориентировочных актов уменьшается. <...>
|
|
|
ГРУППОВОЕ ОБУЧЕНИЕ У МУРАВЬЕВ
Обучение, связанная с ним лабильность поведения и обмен информацией лежат в ос-

Рис. 2. Траектории и этограммы одного из внегнездовых рабочих черноголового муравья:
1 • 24/VII в 9.05, II — в 9.20. Ill — в 18.53, IV—VII — 20/VII в разное время суток; штриховой линией показана фуражировочная дорога
56 Ж.И. Резникова'.
нове механизмов межвидового взаимодействия. В данном разделе мы попытаемся осветить вопросы, необходимые для дальнейшего изложения: каковы изменчивость психических возможностей фуражиров одного и разных видов и способность их к решению сложных задач?
Возможности муравьиного интеллекта давно занимали умы исследователей. Поскольку долгое время господствовало мнение о том, что у насекомых вырабатываются лишь элементарные условные рефлексы, сложность задач, которые экспериментаторы предлагали муравьям, наращивали очень робко. Сама по себе способность муравьев к запоминанию и научению была экспериментально продемонстрирована с помощью различных методик, начиная с классических экспериментов Т. Шнейрлы, обучавшего их в лабиринтах (Schneiria, 1933, 1946), и кончая работами последних лет (Длусский, 1967; Резникова, 1971, 1975; Плеханов, Кауль, 1976; Мазохин-Поршняков, Мурзин, 1976, 1977; Мурзин, 1976, 1977). В частности, при групповом обучении у F . polyctena удалось выработать сложные инструментальные условные рефлексы:
|
|
|
муравьи должны были тянуть за одну из нитей, чтобы получить доступ к кормушке, спрятанной за ширмой. Интересно, что наблюдались случаи, когда тянули за нитку одни муравьи, а проникали за ширму другие, т. е. обучалась именно группа в целом (Rechtsteiner, 1968, 1971).
Приобретение опыта, в том числе основанного на подражании, имеет особенно большое значение для муравьев, так как срок жизни одной особи достигает у некоторых видов 7 лет (Otto, 1962), а в среднем срок жизни рабочих особей — 1,5—2,5 года (Haskins С., Haskins Е., 1980), т. е. больше, чем у многих видов грызунов. В настоящее время считается доказанным существование у муравьев сигнальной преемственности* (Захаров, 1974). При решении задач, требующих объединенных усилий группы особей, или задач, основанных на подражательных реакциях, должна проявляться неоднородность психических возможностей и индивидуального опыта муравьев. Проявление индивидуальных вариаций и способность группы муравьев к решению сложных задач, требующих логических операций, были почти не исследованы до настоящего времени.
|
|
|
Разнокачественность психических возможностей фуражиров. Индивидуальные различия
* Сигнальная преемственность — обучение животного в группе, основанное на подражании другим ее членам (прим. сост.).
в способностях и навыках животных привлекли внимание исследователей в основном в последнее время, когда начались попытки межвидовых количественных сравнений. До сих пор в этом плане внимание уделялось лишь позвоночным животным. Так, Д. М. Рамбо (Rumbaugh, 1968) (цит. по Д. Дьюсбери, 1981) установил, что в группе беличьих обезьян (сай-мири) часть животных решает задачи на "формирование установки" на уровне высших обезьян (более 90% правильных выборов при втором предъявлении), а часть оказывается вообще не способной крещению. К. Н. Благо-склонов (1974) на примере мухоловок-пеструшек, которым предлагались задачи с перемещением гнезд и с преодолением пугающего препятствия (трубка, вставленная в леток), показал, что формы решения задач очень различны и адекватны накопленному индивидуальному опыту.
|
|
|
У муравьев разнообразие стереотипов поведения связано прежде всего с кастовым полиморфизмом и наличием группового и возрастного полиэтизма, т. е. фиксированных различий в функциях, выполняемых разными особями. Этот вопрос довольно хорошо изучен и подробно обсуждался в монографии А. А. Захарова (1972). Однако дифференциация особей может быть еще более глубокой. Известно, что в небольших функционально однородных группах муравьев выделяются "одаренные" особи, которые обладают хорошей памятью и играют роль активаторов при выполнении различных функций и организации групп (Combes, 1941, цит. по А. А. Захарову, 1972).
Различия в способностях и уровне активности рабочих особей можно наблюдать даже в относительно простых ситуациях, когда группа муравьев сталкивается с препятствием на пути к пище или гнезду. Такой опыт был проделан нами в 1968 г. с трофобионтами F . polyctena , которые спускались по стволу березы к муравейнику. Ствол окружили кольцом пластилина с нафталином. Преодоление этого препятствия не было хаотическим: группы из 6—7 фуражиров останавливались перед кольцом и ожидали своего "вожака" — самого активного муравья, который первым преодолевал препятствие и затем пробегал через кольцо туда и обратно, сопровождая остальных муравьев. Возможно, что здесь проявились отношения доминирования-соподчинения, связывающие знакомых между собой особей, которые используют перекрывающиеся поисковые участки.
Поведение муравьев на кормовом участке 57
Предположение о наличии индивидуальной иерархии в группах рабочих особей вытекает и из работы А. М. и Г. Руа (Le Roux А. М., Le Roux G., 1979). В группах из 8—10 особей М. laevinodis наблюдалось постоянное соотношение активно-агрессивных, активно-малоагрессивных, активных и совершенно неактивных муравьев. По этим критериям муравьев перегруппировали в четыре однородные группы. Оказалось, что совместное содержание агрессивных особей приводит к некоторому снижению их агрессивности и активности, а неагрессивных — к появлению среди них агрессивных особей. В результате во всех группах соблюдается примерно то же соотношение особей по уровню агрессивности, что и первичное. При восстановлении первоначальной ситуации проявляется тенденция к сохранению муравьями ранга, приобретенного после первой перегруппировки. Значительное количество муравьев оставались полностью неагрессивными в течение всех перегруппировок.
Сходные результаты получены Г. Верроном (Verron, 1977): одновозрастных особей Lasius niger объединили в четыре группы по уровню активности, которую они проявляли, перетаскивая песчинки. Удаление самых энергичных особей заставляло остальных проявлять большую активность, а их присутствие, напротив, подавляло деятельность остальных.
Эти опыты дают основание полагать, что ранг особей и их поведение в группах зависят от психофизиологических свойств и к тому же поддерживаются активным взаимодействием. Такое взаимодействие было исследовано А. А. Захаровым (1981) в тонком лабораторном эксперименте с малыми группами муравьев Formica rufa .
Оказалось, что индивидуальная борьба за доминирование выражается в увеличении двигательной активности соперничающих особей, а также в проявлениях агрессивности и прямого противоборства. В частности, муравьи устраивают своеобразные турниры, когда претендующий на первенство фуражир пытается занести соперника в гнездо. Два фуражира некоторое время толкают друг друга, стараясь сложить напарника "чемоданчиком". Если это долго не удается ни одному из них, муравьи разбегаются.
Изъятие особей-доминантов из групп со сложившейся поведенческой структурой стимулирует быстрое линейное продвижение оставшихся особей по рангам. При этом выход в лидеры не является формальным перемещением на освободившееся место: именно в пе
риод своего утверждения в новой роли лидер резко увеличивает активность. Вполне возможно, что способность муравьев к обучению коррелирует с их иерархическим положением в группе, обслуживающей определенный участок территории, и с реактивностью по отношению к внутригрупповым ранговым перемещениям.
Пока еще мало данных для того, чтобы сравнивать у разных видов соотношение числа особей, способных быстро справляться с новыми задачами, и особей, не способных к этому. Однако можно полагать, что этот показатель связан со способом использования кормового участка. При коллективной фура-жировке достаточно лишь нескольким особям справиться с задачей, а остальные могут обучаться в результате подражания. Если же муравьи действуют на кормовом участке в одиночку, то существенные различия в их психических возможностях только повлекут за собой неравномерное использование территории: сообразительные особи будут полнее использовать ресурсы своего участка.
Такое предположение подтверждается опытами по коллективному обучению в лабиринтах муравьев нескольких видов. Р. Шовен (Chauvin, 1963) устанавливал лабиринты на фуражировочных дорогах F . polyctena . Это вид с наиболее явно выраженным групповым поведением. В начале опыта только 10—20% муравьев преодолевали лабиринт. Лишь спустя 10 ч, благодаря подражательным реакциям, число их достигло 75—80%.
Аналогичный опыт был проведен нами с F . uralensis в степях Южной Хакасии (1969 г.). Лабиринт, предлагаемый муравьям, представлял собой несколько цилиндров, вставленных один в другой так, чтобы входы в них располагались с противоположных сторон. В последнем внутреннем цилиндре помещалась приманка. До предъявления задачи муравьев прикармливали на площадке для наблюдений в лабиринте первой сложности (один цилиндр). Площадки размещались в периферийной зоне кормового участка. Для того, чтобы исключить возможность накопления муравьями опыта, лабиринты разной сложности расставляли на разных площадках. Оказалось, что в лабиринт второй сложности в течение первого часа наблюдений попадали 70% из пытавшихся проникнуть туда муравьев; за то же время в лабиринт третьей сложности попадали 10—25% особей, а четвертой — всего 1—2%.
Такой же эксперимент, проведенный с Cataglyphis setipes turcomanica (урочище Акибай,
58 Ж. И. Резникова
ТССР, 1970 г.), выявил совсем иное соотношение: 95, 90 и 70% соответственно. У муравьев этого вида кормовой участок организован по принципу одиночной фуражировки, и большинство внегнездовых рабочих легко справлялись с задачей самостоятельно.
Эксперименты, в которых одновременно предъявлялось большое число (50—100) лабиринтов второй сложности, расставленных на всей площади участков наблюдений (1200—1800 м2) в ассоциациях I—Ill, также выявили соответствие между скоростью и характером обучения муравьев и способами организации фуражировки. Для F . picea и F . cunicularia , действующих на территории в одиночку, такая задача вообще не составила сложности, и они доставали приманку при первых же обнаружениях лабиринта. Напротив, среди 20—40 особей F . pratensis , привлеченных к лабиринту, лишь 2—3 муравья проникали к приманке в течение первых четырех часов наблюдений. Впоследствии к ним присоединились еще 8—10 особей, и они вместе транспортировали приманку. Фуражиры М. scabrinodis и L . alienus действовали примерно одинаково: в лабиринте, находящемся вблизи гнезда, появлялись 2—3 разведчика, и в течение 0,5—1 ч они производили. массовую мобилизацию рабочих (до 200 особей). <...>
Задачи, требующие логических операций. Высокий уровень психической организации муравьев вполне позволяет задуматься об их способности усваивать логическую структуру задачи и применять полученный опыт в измененной ситуации. Эти две формы поведения — обучаемость и способность улавливать логические связи — были разграничены Г. Хар-лоу (Harlow, 1958), который таким образом поставил вопрос об объективном изучении рассудочной деятельности животных. По В. Келеру (Kohler, 1921), основной критерий разумного поведения — решение задачи с учетом всей ситуации в целом. По-видимому, именно эта способность муравьев объясняет результаты упоминавшегося выше эксперимента Дж. Брауэра (Brower, 1966), в котором семья Formica Integra , в течение трех лет ежедневно получавшая 10 Р/ч радиации, выстроила крытую дорогу, что позволило уменьшить дозу облучения.
Мнение о принципиальной неспособности насекомых к логическим операциям упорно продолжает сохраняться в научной среде, несмотря на работы Г. А. Мазохина-Поршня-кова (1968, 1969, 1974), посвященные медоносной пчеле. Этот автор экспериментально
доказал, что пчелы могут многое: опознают классы фигур инвариантно к их размеру и взаимному повороту, т. е. обобщают фигуры по форме; способны генерализовать визуальные стимулы по признакам "новизна окраски", "двухцветность", "непарность" (последнюю задачу как наиболее сложную решают лишь единичные особи).
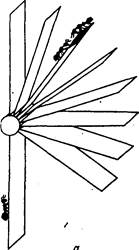

Рис. 3. Экспериментальная установка для исследования группового обучения у муравьев:
а — приманка перемещается в вертикальной плоскости;
б — в горизонтальной
Мы исследовали способность группы муравьев к усвоению логической структуры за-
Поведение муравьев на кормовом участке 59
дачи. Эксперимент проводился с лабораторным гнездом F . polyctena (численность около 1,5 тыс. особей). Здесь уместно сделать небольшое отступление, проливающее некоторый свет на природу экспериментальной работы с муравьями. Дело в том, что в монографии автор обычно старается представить свою работу в виде стройной системы, в которой все эксперименты исторически и логически обусловлены. Однако, как и в некоторых других случаях, опыт, который будет изложен ниже, мы задумали совсем с иной целью: нас интересовало, как муравьям-трофобионтам удается не перепутать ветки с колониями тлей на больших деревьях..
Для изучения ориентации муравьев была сконструирована простая лабораторная установка, схематично изображенная на рис. 3. К вершине стержня (высотой 50 см) прикреплялся веер полосок (каждая длиной 15 см) из плотной бумаги ("ветки"). Одиннадцать "веток" располагались под углом 15° по отношению друг к другу. Приманкой служила капля сахарного сиропа, нанесенного вначале на конец верхней "ветки". Через каждые 10 мин приманка последовательно переносилась с "ветки" на "ветку", все ниже и ниже, помещаясь таким образом под все большим углом к исходному направлению.
Для смены локализации приманки муравьев осторожно снимали с установки, уносили ее с арены, срезали кончик "ветки" с приманкой и, нанеся каплю сиропа на следующую "ветку", вновь предъявляли установку муравьям.
Первые же наблюдения показали, что муравьи, похоже, предлагают нам значительно более интересный вариант опыта (с этим часто приходится сталкиваться): они стали быстро угадывать, на какой "ветке" окажется приманка при очередном предъявлении. Получалось, что группа муравьев усвоила алгоритм решения задачи. Чтобы проверить свое предположение, мы провели серию наблюдений над перемещением приманки и муравьев, учитывая при этом такие параметры, как время, затраченное разведчиками на поиски, доля посещений "правильной ветки" от общего числа посещений, а также скорость движения фуражиров на пути к приманке.
В каждом из 19 опытов (с интервалом 2—3 дня) приманка проходила полный цикл перемещений в вертикальной плоскости (рис. 4, 5). Между 10-м и 11-м опытами был выдержан интервал в 10 дней. Последующие 5 опытов проводились в измененной обстановке: веер "веток" был перемещен в горизонтальную плоскость (рис. 6). Во всех повторностях фиксировали выбор "ветки" каждым приходящим муравьем, а также время, затраченное на поиски пищи первыми тремя особями (дальнейшие поиски в значительной степени оказались обусловлены подражательным поведением). В качестве одного из показателей активности учитывалась скорость движения муравьев по стержню на пути к приманке.
Групповое мечение позволило установить, что приманку посещало в общей сложности около 200 фуражиров. Эта группа сложилась еще в ходе подготовки эксперимента (с непе-
|
|

| 10- |
| Номер "ветки" |
|
|

Рис. 4. Время, затраченное муравьями-первонаходчиками на поиски приманки:
"ветки" расположены в вертикальной плоскости; римские цифры соответствуют номеру опыта
60 Ж. И. Резникова
ремещаемой приманкой), и привлечение новых муравьев было незначительным. Количество муравьев, одновременно посещавших приманку, составляло 30—50 особей. Скорость их движения на пути к приманке возрастала от 8—11 при 1-м предъявлении до 16—20 см за 15 с при 2—3-м предъявлении и далее до конца опыта оставалась на том же уровне. Во время поиска приманки муравьи не ориентировались на искусственный источник света:
включение или перемещение лампы не влияло на скорость обнаружения приманки.
В ходе трех первых опытов время, затраченное разведчиками на поиски приманки,
увеличивалось до 4-го предъявления, т. е. до тех пор, пока направление "ветки" с приманкой не составило 60° по отношению к вертикали. При последующих предъявлениях время поисков сокращалось (см. рис. 4). Причем, если в ходе первых наблюдений муравьи посещали все "ветки", то в дальнейшем — преимущественно те, на которых они получали приманку при более ранних предъявлениях. Основная доля посещений приходилась на ту "ветку", где приманка была 10 мин назад. Таким образом, поиски приманки постепенно упорядочивались: муравьи запоминали ее расположение, но не делали поправки на перемещение (см. рис. 5, а).
Дата добавления: 2019-07-15; просмотров: 174; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!