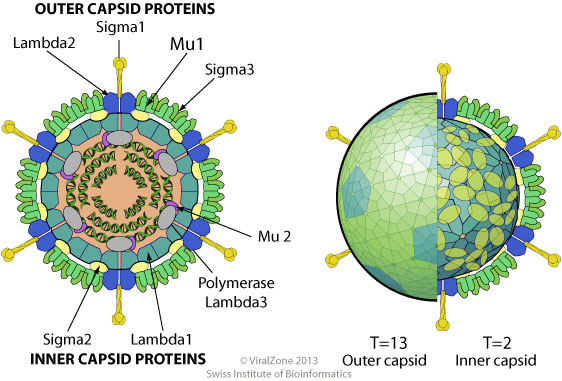
Расположение белков MRV в вирионе
Проникновение ортореовирусов в клетку
Ø После присоединения к рецепторам (молекулы, содержащие сиаловые кислоты), ортореовирусы интернализуются в эндосомы.
Ø В эндосомах происходит протеолиз δ3 и μ1С и образуется ISVP (промежуточная вирусная частица).
Ø ISVP проникает в цитоплазму, где происходит окончательное «раздевание» до кора.
Синтез мРНК ортореовирусов
Ø Синтез мРНК происходит внутри коровых частиц.
Ø С каждого сегмента транскрибируется полноразмерная мРНК, которая кэпируется, поли(А) отсутствует.
Ø Синтезированные мРНК транспортируются из кора через «башни» в вершинах. Родительская дцРНК при этом остаётся внутри
Ø Есть предположение, что каждая «башня» – это независимая транскрипционная единица для «своего» сегмента дцРНК
Трансляция белков MRV
Ø 10 мРНК транслируются в 12 белков
Ø С мРНК сегмента μ3 транслируется 2 белка с 2х AUG-кодонов, находящихся в рамке: μNS и (μNSC)
Ø С мРНК сегмента S1 транслируется 2 белка с AUG-кодонов, находящихся не в рамке. Соответственно, образуется 2 разных белка: δ1 и δ1NS
Ø Из 12 белков МRV 4 – неструктурные, а 8 являются компонентами вириона.
Сборка вирионов MRV
Ø мРНК MRV служат интермедиатами для репликации реовирусов. После выхода из кора мРНК связываются с неструктурными белками μNS,δ1NS и δЗ. Далее к этим комплексам присоединяются λ2 и λ3 (РНК–полимераза), после чего происходит синтез второй цепи РНК.
|
|
|
Ø Во время синтеза дцРНК происходит формирование вирионов.
Ø дцРНК всегда находится внутри частиц для предотвращения распознавания клеточными молекулярными сенсорами.
Ø Белок μ1 во время созревания расщепляется на 2 белка: μ1C и μ1N
Жизненный цикл реовирусов
Вирионы реовирусов могут адсорбироваться (с помощью белка δ-1) на клетке и проникать рецептор-опосредованным эндоцитозом в цитоплазму, где под влиянием ферментов лизосом происходит частичная депротеинизация. Возможно проникновение вирусов в клетку другим механизмом, например, инфекционных субвирусных частиц (ИСЧ), не содержащих белка δ-1. ИСЧ ротавирусов проникают через клеточную мембрану; механизм проникновения неизвестен. ИСЧ освобождают сердцевину в цитоплазме, и ферменты сердцевины инициируют продукцию иРНК. С каждого фрагмента геномной РНК считывается индивидуальная иРНК. Транскрипция генома проходит в две фазы (ранняя и поздняя). Минус нить РНК используется как матрица. Сборка вирионов происходит в цитоплазме. Вирусы выходят при лизисе клетки.
Ротавирусная инфекция
Ø Вызывает инфекционную диарею, в основном у новорождённых и детей
|
|
|
Ø В развивающихся странах диарея (около 50% которые вызывается ротавирусом) – основная причина детской смертности
Ø По разным оценкам в мире от ротавирусной диареи погибает 300000– 800000 человек в год
Вирион ротавирусов сферический (70 нм). Геномная дц РНК (18 550н.п.) имеет содержание A+U на уровне 58-67%. В вирионах не содержатся олигонуклеотиды. Трехслойный капсид окружает 11 сегментов генома (от 667 до 3 302 н.п.), два из которых (9-й и 11-й сегменты) имеют по две открытые рамки считывания (ORF). Из 13 вирусных белков три гликозилированы (в VP7 и NSP4 – аспарагин, а в NSP5 – серин и/или треонин). Формирующийся вирион во время почкования незрелых частиц через систему эндоплазматического ретикулума клетки временно приобретает дополнительную липидную оболочку, которая утрачивается в зрелой частице
На основании перекрестных РТГА выделяют 7 серотипов: А, В, С, D, Е, F, G. Ротавирусы серотипов А, В и С обнаружены у человека и животных; D, Е, F, G – только у животных.
Жизненный цикл ротавирусов
Вирионы могут проникать рецептор-опосредованным эндоцитозом в клетку , где под влиянием ферментов лизосом происходит частичная депротеинизация — разрушение наружного капсида с образованием субвирусных частиц. Однако это тупик для ротавирусов. Другой механизм проникновения заключается в том, что вирионы ротавирусов активируются протеазами (например, в ЖКТ), превращаясь в инфекционные субвирусные частицы, которые пенетрируют клеточную мембрану и в цитоплазме утрачивают наружный капсид (под действием лизосом), освобождая сердцевину . Ферменты сердцевины инициируют продукцию иРНК, используя в качестве матрицы минус-нить РНК. Белки VP7 и NS28 синтезируются как гликопротеины и экспрессируются в эндоплазматическом ретикулуме ЭР. Плюс-РНК является иРНК. Она включена внутрь капсидов как матрица для репликации +/- сегментированного генома. Капсиды ротавирусов агрегируют, связываются с белком NS28 в (ЭР) и приобретают белок VP7 наружного капсида. Вирусы выходят при лизисе клетки.
|
|
|
Антиротавирусные вакцины
Ø Существует 4 основных серотипа ротавируса А (90% всех случаев), поэтому все разрабатываемые вакцины квадривалентны.
Ø В 90-е гг. была лицензирована вакцина RotaShield – живая вакцина на основе ротавируса макаки-резуса
Ø К сожалению у этой вакцины на каждые 2000 применений случаются серьёзные осложнения – непроходимость кишечника
|
|
|
Ø Сейчас лицензированы 2 новые вакцины: Rotateq (Merck) и Rotarix (GlaxoSmithKline)
17. Ретровирусы. Обратная транскрипция и интеграция в геном клетки-хозяина. Вирус Т-клеточного лейкоза человека (HTLV/PTLV). Заболевания, ассоциированные с HTLV, географическая распространённость. Вирус иммунодефицита человека (ВИЧ/HIV). Изменчивость ВИЧ. СПИД, основные клинические проявления. Терапевтические средства.
Ретровирусы
Ретровирусы — обширное семейство оболочечных вирусов, имеющих PНK-геном, — отличаются от прочих РНК-содержащих вирусов способностью осуществлять обратную транскрипцию вирусной РНК с образованием ДНК и интеграцией её в геном хозяйской клетки. Характерной особенностью этой группы вирусов является очень высокая степень изменчивости, которая связана с большой частотой ошибок, происходящих в ходе обратной транскрипции. Все ретровирусы обладают сходной морфологией вириона и организацией структуры генома, однако различия их патогенных свойств весьма велики — некоторые вирусы этой группы вообще непатогенны для своих хозяев.
Ø Семейство: Retroviridae
- подсемейство: Orthoretrovirinae
- род: Deltavirus (HTLV-1, HTLV-2)
- род: Lentivirus (HIV-1, HIV-2)
- подсемейство: Spumaretrovirinae
Линейная ssRNA (димер), 7-11 kbp, d=80-100 нм, внешняя липопротеиновая оболочка
В жизненном цикле ретровирус обязательно проходит стадии обратной транскрипции и встраивания в геном хозяина.

У всех ретровирусов в геноме закодировано как минимум 3 гена: gag, pol и env
5'-сар, З'-polyA, на 5'-конце находится PBS (сайт посадки праймера), а на З'-конце - РРТ (полипуриновый тракт)
LTR (Long terminal repeat: U3, R, U5)
Белки группы Gag (group specific antigens) формируют внутреннюю структуру вириона и включают три основных полипептида: капсид (СР), ответственный за пространственную укладку молекул всех прочих белков и формирование архитектуры вириона; матриксный белок (МА), тесно связанный с внутренней поверхностью липидного бислоя и стабилизирующий вирусную частицу; нуклеокапсидный белок (NC), находящийся в тесной связи с вирусной РНК и обеспечивающий её конденсацию внутри нуклеокапсида. Помимо этого, группу Gag некоторых ретровирусов дополняют другие белки, имеющие общий с основными белками предшественник и принимающие участие в формировании и почковании вирионов.
Ферменты, ответственные за различные этапы размножения ретровирусов, составляют вторую группу белков — Pol, включая протеазу (Pro), обратную транскриптазу (RT) и интегралу (Int).
Третья группа ретровирусных белков — Env — включает два гликопротеина: поверхностный (SU) и трансмембранный (ТМ), формой напоминающий «шипы» и «пронизывающий» вирусную мембрану (в составе вириона SU и ТМ образуют «тройки» — тримеры). Белок SU ответственен за связывание вириона с рецепторами, а ТМ обеспечивает слияние мембран в ходе проникновения вируса в чувствительные клетки.
Интеграция
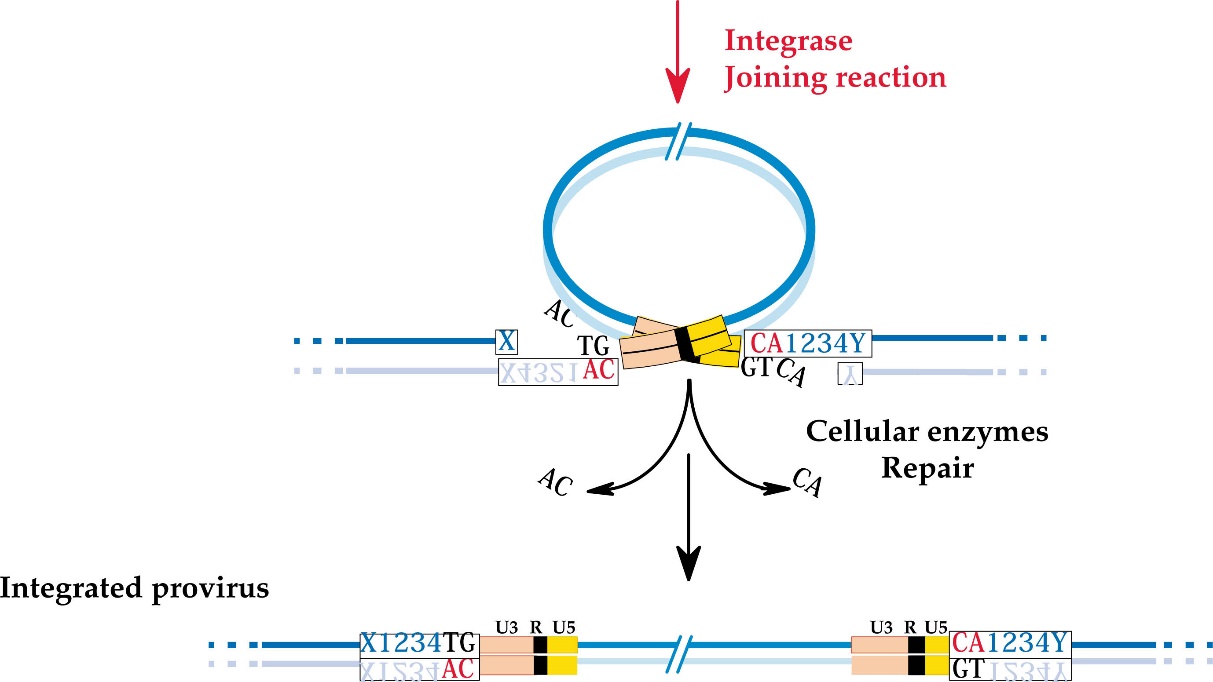
Геномы ретровирусов обнаруживаются в интегрированном провирусном состоянии в зараженных и трансформированных клетках. РНК-геном ретровирусов подвергается обратной транскрипции с образованием ДНК на начальном этапе репликативного цикла, после чего образованная ДНК (провирус) интегрирует с геномом клетки-хозяина. Механизм интеграции ретровирусов имеет три отличительные особенности: а) место включения в хозяйскую хромосому неспецифично и не имеет гомологии с вирусными последовательностями, что указывает на интеграцию посредством незаконной рекомбинации; б) короткая нуклеотидная последовательность в участке интеграции дуплицируется, что приводит к образованию прямых повторов нескольких оснований ДНК хозяина, фланкирующих провирус; в) участок интеграции в вирусной ДНК строго детерминирован. Таким образом, интеграция ретровирусов происходит путем встраивания специфического сайта вирусной ДНК в неспецифический сайт клеточной ДНК. Способность ретровирусов захватывать клеточные гены и включать их в хозяйскую хромосому приводит к мысли о том, что, возможно, эти вирусы играют роль агентов, переносящих генетическую информацию наподобие трансдуцирующих фагов.
Транскрипция ретровирусов
Транскрипция осуществляется клеточной РНК-полимеразой II узнающей необходимые сайты инициации на 5’LTR. Кэпирование и полиаденилирование также происходит за счет хозяйских ферментов. Часть вирусных РНК-транскриптов экспортируется из ядра в цитоплазму в несплайсированном виде. У одних ретровирусов в РНК присутствуют специальные последовательности (~150 нуклеотидов), которые связываются с хозяйскими факторами, обеспечивающими экспорт из ядра. Другие ретровирусы (HTLV, HIV) кодируют специальные белки, которые транспортируют непроцессированную РНК в цитоплазму.
Дата добавления: 2018-06-01; просмотров: 453; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!