Симптоматика гемморагических лихорадок Марбург и Эбола
Ø Инкубационный период 2-21 день (в среднем 4-10)
Ø Начальная фаза: неспецифические симптомы – лихорадка, озноб и общее недомогание
Ø Далее: прострация, анорексия, рвота, боли в груди и одышка
Ø Пик инфекции: нарушение работы сосудов (проявляющееся в эритеме), спонтанные кровотечения (массивные кровопотери редки), далее – диффузнаякоагулопатия
Ø Летальные случаи заканчиваются шоком, множественным поражением внутренних органов, комой и смертью в течение 6-16 дней
Ø Выжившие имеют серьезные нарушения иммунной системы, подвержены глазным болезням, миалгии, миелиту; часто развиваются психозы
Патогенез филовирусных геморрагических лихорадок
Ø Главная причина смерти –нарушение сосудистой системы организма
Ø 3 основных процесса:
1. Увеличение проницаемости сосудов
2. Диссеминированное внутрисосудистое свертывание (ДВС-синдром)
3. Нарушения в иммунном ответе
Патогенез филовирусных геморрагических лихорадок
Ø Инфекция моноцитов/макрофагов ведет к экспрессии провоспалительных цитокинов (IL-6,TNF-α, IL-1β)
Ø Инфекция дендритных клеток ведет к ингибированию синтеза молекул MHC-I/II, CD80, CD86, CD40
Ø TNFαи NO увеличивает проницаемость сосудов и усиливают экспрессию поверхностных молекул адгезии на эндотелии
Ø Один из основных факторов патогенеза – тромбопластин (TF, фактор III). Активно продуцируется инфицированными макрофагами и клетками эндотелия (после воздействия TNFα)
|
|
|
Ø Высокая скорость репликации вируса – важный фактор патогенеза филовирусной инфекции. Вскоре после начала заболевания в различных органах, в особенности в печени и селезенке, обнаруживается высокий титр вируса
Ø Блокировка вирусом IFN-I-ответа способствует быстрому распространению инфекции по организму. Индукцию интерферонов α- и β- блокирует VP24 филовирусов (+VP35 ZEBOV)
Лечение филовирусных геморрагических лихорадок
Ø Обезболивающие препараты
Ø Применение сыворотки выздоровевших больных (имело ограниченный успех)
Ø Рекомбинантный антикоагулирующий белок нематоды с2 (rNAPc2)
Ø Антицитокиновая терапия
Ø siRNA, короткие пептиды
Вакцины против филовирусов
Ø На данный момент одобренной к применению на людях вакцины не существует
Ø Целый ряд вакцин был эффективен на моделях грызунов, но провалился в экспериментах на приматах
Ø Нереплицирующиеся аденовирусы, экспрессирующиеGP и NP (ZEBOV)
Ø Рекомбинантные VSV, экспрессирующие 1) ZEBOVGP 2)MARVGP (показали эффективность и в терапии)
Ø ВВЭЛ, экспрессирующийMARVGP и/или NP
16. Реовирусы: классификация, структура вириона, строение генома. Жизненный цикл. Функции белков. Ротавирусная инфекция.
|
|
|
Реовирусы
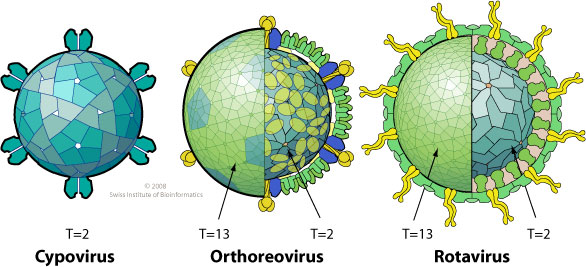
Ø сегментированная линейная dsRNA; 10,11 или 12 сегментов общей длиной 16-27 kb
Ø капсиды, как правило, двойные или даже тройные (исключение: Ципо- и Диновернавирусы), также наружный капсид у некоторых родов может быть пронизан широкими «башенподобными» белковыми шипами; d=60-80 нм
Ø очень большое семейство – на сегодняшний день описано уже 15 родов
Ø хозяева: позвоночные, беспозвоночные, растения, грибы
Положительная цепь в каждом дуплексе РНК кэпирована по 5'-концу; 3'- и 5'-концы плюс-цепи РНК содержат короткие консервативные последовательности, участвующие в процессах транскрипции и репликации вирусных РНК. Обе нити РНК терминированы 3'-ОН без полиаденилирования.
Белки внутреннего капсидного слоя обеспечивают репликацию геномных РНК и формирование мРНК, а структурные белки среднего и/или наружного слоя капсида защищают вирусный геном и обладают антигенной активностью. По крайней мере, 3 внутренних белка обладают энзиматической активностью, связанной с репликацией РНК и кэпированием: наиболее консервативная РНК-зависимая РНК-полимераза (роl), которая может функционировать и как транскриптаза, и репликаза; нуклеоnидфосфогидролаза; гуанилилтрансфераза; две метилтрансферазные активности и хеликаза. Несколько минорных белков выполняют структурную роль вместе с тремя (по крайней мере) капсидными белками.
|
|
|
Капсидные белки реовирусов позвоночных обладают видоспецифической и типоспецифической антигенной активностью. У вирусов беспозовночных и растений белки более униформны и менее вариабельны как АГ.
Классификация реовирусов

Между вирусами отдельных родов нет антигенных перекрестов. Некоторые вирусы вызывают гемагглютинацию.
Число суперкапсидных слоев является отличительной чертой каждого вида реовирусов. По этому их можно разделить на 2 группы:
Ø вирусы с шипами на поверхности капсида или зрелого вириона в 12 вершинах икосаэрических фасеток (Ortoreovirus, Cypovirus, Aquareovirus, Oryzavirus, Fijurus, Idnovirus, Mycovirales)
Ø вирусы с относительно «гладкой поверхностью», без выступов и шипов (Orbivirus, Rotavirus, Seadornavirus, Coltivirus, Phytoviruses)
Важные патогенны реовирусов
| Род/вид | Хозяин | Трансмиссия/вектор | Заболевание | ||
| Ortoreovirus | |||||
| Реовирус млекопитающих(MRV) | Человек, овца, КРС, свиньи | Орально-фекальный | Гастроэнтерит, респираторные заболевания | ||
| Orbivirus
| |||||
| «Синего языка»(BTV) | Овцы, КРС | Мокрецы рода Culicoldts | Ринит, стоматит | ||
| Чингинола(CGLV) | Человек | Москиты | Лихорадка | ||
| Кемерово | Человек | Клещи | Лихорадка, энцефалит | ||
| Coltivirus | |||||
| Колорадская клещевая лихорадка(CTPV) | Человек | Клещи | Лихорадка, энцефалит | ||
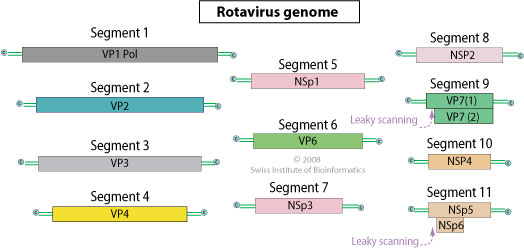
| Rotavirus | |||||
| Группа А(RV-A) | Человек, животные | Орально-фекальный | Детская диарея | ||
| Группа В(RV-B) | Человек, животные | Орально-фекальный | Эпидемия диареи взрослых | ||
| Группа С(RV-C) | Человек, животные | Орально-фекальный | |||
| Seadomavirus | |||||
| Банна(BAV) | Человек | Комары рода Culex | Энцефалит | ||
Геном ортореовирусов
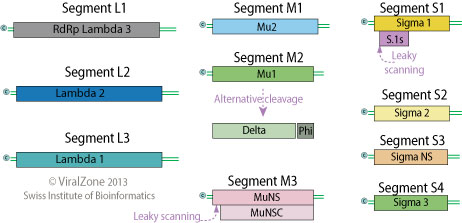
Ø 12 «шипов», отходящих от поверхности нуклеокапсида в вершинах икосаэдрических фасеток, 10 сегментов дцРНК (3 больших (L), 3 средних (М) и 4 малых (S), кодирующих соответственно 3 λ-, 3 μ- и 4δ-белка), причём 10-й полицистронный сегмент кодирует более и мелкие генные продукты. δ-1-белок является серотип-специфическим антигеном (10 сегментов, 23.5 kb)
Ø За счёт сегментации реовирусы могут осуществлять реассортацию (при коинфекции происходит обмен фрагментами генома (только между близкородственными вирусами))
Ø 3'- и 5'-концы высококонсервативны
Ø Большая часть белков реовирусов (в т.ч. ферментативный аппарат) является компонентами вириона
В центре вириона располагается РНП (Рибонуклеопротеиды – состоят только из вирусной РНК и специфических вирусных белков) диаметром 48 нм, составленный из геномной дцРНК, окружённой 120 копиями белка «λ-1» (HEL). Снаружи РНП располагается внутренний капсид диаметром 60 нм.
Белки ортореовирусов
| Сегмент | Белок | Длинна сегмента (нукл.) | Функции белка | Локализация/молекул на вирион |
| L1 | λ3 | 3854 | РНК-полимераза | Кор/12 |
| L2 | λ2 | 3916 | Гуанилил трансфераза | Башни на коре/60 |
| L3 | λ1 | 3896 | НТФаза, хеликаза | Кор/120 |
| M 1 | μ2 | 2304 | Кофактор полимеразы? | Минорный компонент кора/12 |
| M2 | μ1→ μ1C | 2203 | Основной структурный белок | Внешняя капсидная оболочка/600 |
| M3 | μNS(μNSC) | 2235 | одРНК-связывающий белок | Неструктурный белок |
| S1 | δ1 | 1416 | Гемагглютинин, рецепторный белок | Капсид/36 |
| δ1NS | ? | Неструктурный белок | ||
| S2 | δ2 | 1331 | Структурный белок | Кор/240 |
| S3 | δNS | 1198 | одРНК-связывающий белок | Неструктурный белок |
| S4 | δ3 | 1196 | Основной структурный белок | Внешняя капсидная оболочка/600 |

У ротавирусов внутренний капсид включает белки VP1, VP2, VP3, VP6. Наружный капсид реовирусов состоит из белков δ-1, δ-3, μ-1С, а также белков λ-2, отходящих от сердцевины и выступающих в виде шипов. Белок δ-1 является гемагглютинином и прикрепительным белком. Белок μ-1С определяет способность реовирусов заражать клетки кишечника и впоследствии поражать ЦНС. У ротавирусов наружный капсид включает белки VP4 (шипы, выступающие на поверхности вириона, являющиеся гемагглютинином и прикрепительным белком) и VP7 – основной компонент наружного капсида, являющийся типоспецифическим АГ. Ротавирусы и ортореовирусы активизируются протеолизом (инфекционные субвирусные частицы — ИСЧ) с увеличением их инвазионной способности.
Дата добавления: 2018-06-01; просмотров: 395; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!