Элементы сравнительной эмбриологии позвоночных
Закон зародышевого сходства Бэра и его современная трактовка.
Бэр продемонстрировал единство плана строения зародышей различных классов позвоночных. Это привело его к важнейшему эмпирическому обобщению - «закону зародышевого сходства» (начало XIX в). Бэр утверждал следующее: зародыши различных видов, относящихся к одному типу, более сходны между собой, нежели взрослые формы, и их видовые различия нарастают по ходу развития. Иными словами, сначала в развитии появляются черты типа, потом класса и т. д.
В основе действия закона зародышевого сходства лежит большая жизнеспособность тех мутантов, у которых фенотипический эффект мутаций проявляется на более поздних стадиях онтогенеза; рано проявляющиеся мутации чаще приводят к нарушениям работы сложных корреляционных систем в развивающемся организме, что ведёт к гибели зародыша. Поэтому онтогенез в целом проявляет тенденцию оставаться консервативным (особенно на ранних стадиях). 3ародышевое сходство разных видов есть следствие их филогенетического родства и указывает на общность происхождения, что впервые подчеркнул Ч. Дарвин.
Выше говорилось о ЗЗС, сформулированного Бэром на основе сопоставления хода развития различных классов позвоночных: чем раньше стадия развития, тем более сходной является структура зародышей, относящихся к различным систематическим группам. Это означает, что, по Бэру, в ходе развития нарастает дивергенция (расхождение) признаков. Оценивая сегодня этот закон следует учесть, что Бэр не был знаком с более ранними, чем нейрула, стадиями развития позвоночных. Исходя из полученных позже сведений картина рисуется иначе: как раз наиболее ранние стадии развития - дробление, гаструляция - в развитии различных классов позвоночных могут проходить совершенно по-разному: достаточно сопоставить гаструляцию у костистых рыб, амфибий и амниот. На поздних стадиях развития также, конечно, имеются существенные систематические различия: применительно к этим стадиям закон Бэра полностью сохраняет свою силу. Однако, имеется период развития, который можно назвать «узлом сходства», поскольку в этот период у всех без исключения позвоночных сходным путем развиваются сходные структуры: это период закладки осевых органов - нервной трубки, хорды, сомитов и других сопутствующих образований. Зародыши различных классов позвоночных различными путями (например, проходя или не проходя через инвагинационную гаструляцию) подходят к этому этапу, но затем проходят его весьма единообразно. Иногда данную стадию развития называют «фарингула» (от греческого pharynx - глотка).
|
|
|
 Такое единообразие связано с тем, что в основе нейруляции и закладок осевых органов у всех позвоночных лежит одно и то же в принципе морфогенетическое движение: передне-заднее растяжение - латеромедиальная конвергенция материала дорсальной эктодремы и мезодермы. Именно это движение, лежащее в основе формирования центральной нервной системы и сопутствующих ей двигательных и опорных органов, и определяет единый тип развития и строения всех позвоночных.
Такое единообразие связано с тем, что в основе нейруляции и закладок осевых органов у всех позвоночных лежит одно и то же в принципе морфогенетическое движение: передне-заднее растяжение - латеромедиальная конвергенция материала дорсальной эктодремы и мезодермы. Именно это движение, лежащее в основе формирования центральной нервной системы и сопутствующих ей двигательных и опорных органов, и определяет единый тип развития и строения всех позвоночных.
|
|
|
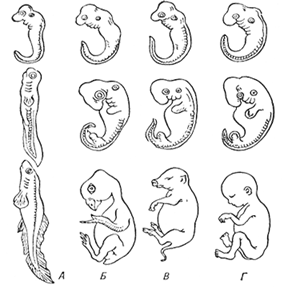
Последовательные стадии развития зародышей рыбы (Л), курицы (Б), свиньи (В), человека (Г)
Морфогенетические движения в раннем развитии костистых рыб.
Яйцеклетки с большим количеством желтка, который распределен неравномерно вдоль анимально-вегетативной оси яйца (полилецитальные, резко телолецитальные яйцеклетки), претерпевают неполное дискоидальное дробление. Так дробятся зиготы костистых рыб, рептилий и птиц. В результате такого дробления образуется зародыш - дискобластула. Дискобластула характеризуется следующими признаками:
1) форма сферическая;
2) крыша бластулы имеет вид небольшого многоклеточного диска, называемого бластодиском, или бластодермой, а дно представлено объемной массой нераздробившегося желтка;
|
|
|
3) между бластодиском и желтком имеется узкое пространство подзародышевая полость (щелевидная).
Дробление костистых рыб. В яйцеклетках костистых рыб после оплодотворения начинается движение цитоплазмы по направлению к анимальному полюсу, где образуется скопление свободной от желтка цитоплазмы - будущий бластодиск. Масса недробящегося желтка называется желточной клеткой. По периферии бластодиска его цитоплазма непосредственно переходит в тонкий поверхностный слой цитоплазмы желточной клетки - желточный цитоплазматический слой. Такая сегрегация цитоплазмы от остального объема яйцеклетки, занятой желтком, продолжается в течение нескольких делений дробления.
Дискоидальное дробление у костистых рыб характеризуется тем, что его первые 4-5 борозд проходят меридионально (вертикально) и под прямым углом друг к другу. Дискоидальное дробление, как и полное неравномерное дробление, полностью отвечает правилам Гертвига-Сакса.
Борозда первого деления дробления закладывается на анимальном полюсе и распространяется вертикально по направлению к вегетативному, но разделяет только цитоплазму бластодиска, так что образовавшиеся бластомеры снизу и по периферии бластодиска не отграничены от желтка. Борозда первого деления проходит поперек длинной оси бластодиска, который имеет эллипсоидную форму при взгляде со стороны анимального полюса. Борозда второго деления дробления проходит перпендикулярно первой борозде вдоль длинной оси бластодиска. Борозды третьего деления параллельны борозде первого, а борозды четвертого деления параллельны бороздам второго.
|
|
|
На стадии 16 бластомеров впервые появляются клетки, полностью отделенные друг от друга и от желтка, - это квартет самых центральных бластомеров. Такое полное отделение происходит в конце четвертого цикла, когда борозды деления отсекают бластодиск от центра, распространяясь к краю бластодиска. Периферические 12 бластомеров называют краевыми бластомерами. Их цитоплазма переходит непосредственно в желточный цитоплазматический слой.
Борозда пятого или шестого деления дробления проходит горизонтально (параллельно экватору яйца), в результате этого образуется второй, внутренний ярус бластомеров, называемых глубокими клетками. Из глубоких клеток образуется непосредственно зародыш костистых рыб. Каждая глубокая клетка является одной из двух дочерних клеток четырех центральных бластомеров на 32-клеточной стадии. Другие дочерние клетки остаются поверхностными. Дочерние клетки краевых бластомеров также становятся поверхностными.
В последующем появляются тангенциальные борозды, при этом направление борозд дробления подвержено большим индивидуальным колебаниям. Деления дробления становятся «метасинхронными», поскольку в ходе каждого клеточного цикла по бластодиску проходит волна митозов. Клетки на анимальном полюсе вступают в митоз первыми, краевые бластомеры последними. Зародыш на стадии 256 бластомеров представляет собой морулу, где бластомеры образуют бугристую шапочку, возвышающуюся над желтком.
После 9-11 деления клеточный цикл удлиняется, наступает десинхронизация делений, появляется подвижность глубоких бластомеров.
Краевые бластомеры ранней бластулы имеют уникальную судьбу. Начиная с 10-го клеточного цикла, боковые мембраны этих влеток исчезают, а цитоплазма и ядра сливаются с желточным цитоплазматическим слоем желточной клетки. Так образуется желточный синцитиальный слой с одним рядом ядер в виде узкого кольца вокруг края бластодермы. После образования желточного синцитиального слоя клетки вышележащего яруса поверхностных бластомеро встановятся краевыми, но в отличие от краевых бластомеров предыдущих стадий они полностью отделены мембранами от желточной клетки. Ядра желточного синцития претерпевают 3-5 метасинхронных делений, количество рядов ядер к стадии средней бластулы возрастает. К поздней бластуле часть ядер синцития перемещается под бластодерму, образуя внутренний желточный синцитиальный слой, другая часть желточного синцития спереди от края бластодермы формирует наружный желточный синцитиальный слой. Часть ядер внутреннего слоя может образовываться за счет деления самых базальных глубоких бластомеров. Желточный синцитиальный слой непосредственно не участвует в построении тела зародыша. Это экстраэмбриональное образование, имеющее важное морфогенетическое занчение и обеспечивающее эпиболию бластодермы во время гаструляции.
Поверхностные бластомеры в процессе бластуляции образуют эпителиальный монослой бластодермы - покровный слой. Вначале поверхностные бластомеры делятся как в плоскости пласта, так и перепендикулярно ему. Планарные деления дают дочерние клетки, отсающиеся в покровном слое, а при перпендикулярном делении одна дочерняя клетка остается в покровном слое, другая же становится глубокой клеткой. По мере развития клетки покровного слоя сильно уплощаются, покровный слой становится трудно различимым. Боковые поверхности бластомеров покровного слоя образуют плотные и промежуточные контакты, которые обеспечивают изоляцию внутренней среды зародыша от окружающей среды. Краевые бластомеры покровного слоя и прилежащие к ним участки желточного синцитиального слоя соединены плотным и следующим за ним протяженным промежуточным контактом. К стадии поздней бластулы деления бластомеров покровного слоя совершаются только в плоскости пласта, а после начала эпиболии и до ее конца число клеток покровного слоя не меняется. Клетки покровного слоя не участвую в построении тела зародыша.
Таким образом, к стадии поздней бластулы в зародышах костистых рыб формируются три морфологически обособленных клеточных компартмента с разной судьбой - глубокие клетки (эпибласт), поверхностные клетки (перидерма) и желточный синцитиальный слой (перибласт). Трехслойная структура бластодермы - особенность, общая для всех костистых рыб.
Желточный синцитий играет ведущую роль в эпиболии: его край активно сокращается (благодаря эндоцитозу участков клеточных поверхностей) и подтягивает в вегетативном направлении слой поверхностных клеток, которые при этом перегруппировываются. По завершении эпиболии образуется желточный мешок, который впоследствии втягивается внутрь зародыша, входя в состав его кишечника. Однако еще много раньше (когда обрастание охватило не более половины поверхности яйцеклетки) в той части бластодиска, которая будет соответствовать дорсальной стороне зародыша, протекают основные формообразовательные процессы. Важное отличие костистых рыб от осетровых и амфибий состоит в том, что у первых отсутствует типичное гаструляционное подворачивание (инволюция). Она заменена ингрессией - вселением под поверхность бластодиска небольшой группы клеток. Основную роль в формировании осевых зачатков у костистых рыб играют уже знакомые нам по развитию амфибий движения конвергентной интеркаляции клеток к дорсальной средней линии (рис. 58, А, Б). При этом все осевые зачатки (хорда, сомиты, нервная трубка) образуются из внутренней массы бластомеров, с самого начала лежащих на разных уровнях. Это означает, что карта презумптивных зачатков у костистых рыб с самого начала является трехмерной (рис. 58, В). Клетки наружного слоя образуют лишь внешнюю часть эктодермы зародыша. Интересно, что у карпозубой рыбы нотобранха, обитающей в пересыхающих водоемах тропических стран, поверхностные бластомеры вовсе не участвуют в развитии зародыша, а образуют плотную оболочку, предохраняющую яйца от высыхания. Данное приспособление можно рассматривать.ю как аналог амниотических оболочек высших позвоночных.
П  ри нейруляции у костистых рыб не происходит типичных для большинства зародышей позвоночных движений скручивания нервной пластинки. Вместо этого презумптивыне клетки нервной системы конвергируют к средне-дорсальной линии зародыша, одновременно с другими клетками осевых зачатков, а невроцель возникает в этой клеточной массе шизоцельным путем (т.е. путем расхождения клеток).
ри нейруляции у костистых рыб не происходит типичных для большинства зародышей позвоночных движений скручивания нервной пластинки. Вместо этого презумптивыне клетки нервной системы конвергируют к средне-дорсальной линии зародыша, одновременно с другими клетками осевых зачатков, а невроцель возникает в этой клеточной массе шизоцельным путем (т.е. путем расхождения клеток).
Дата добавления: 2018-05-09; просмотров: 831; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!