Гемоглобиновая буферная система
В физиологическом диапазоне рН оксигемоглобин является более сильной кислотой, чем восстановленный гемоглобин. Поэтому, освобождая в тканях кислород, восстановленный гемоглобин приобретает большую способность к связыванию ионов водорода.
Гемоглобиновый буфер обеспечивает 35% буферной емкости крови.
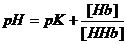
Гемоглобин является амфотерным веществом. В тканях редуцированный гемоглобин проявляет основные свойства:

Оксигемоглобин, образующийся в легких, ведет себя как кислота, поэтому смещения рН в кислую сторону не происходит. В капиллярах НbО2 теряет часть кислотных свойств. Образующийся редуцированный гемоглобин связывает ионы водорода и освобождает ионы калия, которые при ацидозе выходят из эритроцитов, вызывают гиперкалиемию. Ионы калия затем выводятся почками.
Под влиянием избытка СО2 сродство гемоглобина к кислороду падает, а диссоциация оксигемоглобина повышается. Уменьшение емкости гемоглобинового буфера происходит при кровопотере, гемолизе, анемиях.
Концентрация водородных ионов почти полностью определяется соотношением бикарбоната и углекислоты. Содержание этих веществ в крови тесно связано с процессом переноса кровью углекислого газа (СО2) от тканей к легким. Физически растворенный СО2 диффундирует из тканей в эритроцит в соответствии с градиентом концентрации, поскольку в эритроцитах образуется мало углекислого газа. В эритроците под действием фермента карбоангидразы происходит гидратация молекулы СО2 с образованием угольной кислоты (Н2СО3), сразу же диссоциируя на ион бикарбоната и ион водорода:
|
|
|
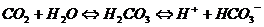
Накапливающийся ион НСО3ˉ диффундирует по градиенту концентрации в плазму. Электрохимическая нейтральность поддерживается за счет перемещения в эритроциты ионов хлора (хлоридный сдвиг).
В физиологических условиях повышение рСО2 в венозной крови стимулирует образование НСО3ˉ в эритроцитах. Наоборот, снижение рСО2 в артериальной крови угнетает образование бикарбоната. При этом обеспечивается постоянство артерио-венозной разницы НСО3ˉ/СО2 и, следовательно, рН.
При патологических процессах, сопровождающихся изменениями рСО2, эти механизмы сохраняются. Повышение рСО2 в эритроцитах способствует увеличению внеклеточного бикарбоната, и наоборот.
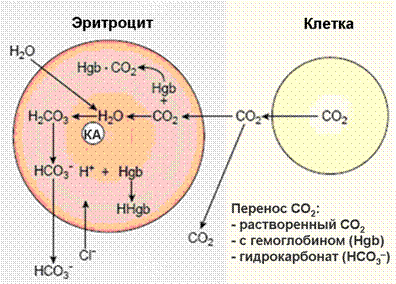
Рис.4.Химические реакции, протекающие в эритроцитах при газообмене в тканях.
Буферные свойства крови обусловлены, в конечном итоге, суммарным эффектом всех анионных групп слабых кислот, важнейшими из которых являются бикарбонаты и анионные группы белков. Эти анионы, обладающие буферными эффектами, получили названия «буферные основания» (buffer bases, BB).
|
|
|
Клетки являются постоянным источником углекислого газа и нелетучих кислот. Часть ионов водорода способна через метаболически активные механизмы поступать в клетки в обмен на ионы калия. В клеточном секторе преимущественное значение имеют фосфатная и белковая буферные системы. Буферные основания представлены в основном калийными солями фосфорной кислоты и белков.
Калиево-водородный обмен в плазме.
Свободное перемещение одноименно заряженных ионов калия (К+) и водорода (Н+) между внутриклеточным и внеклеточным секторами организма имеет важное значение в регуляции кислотно-основного равновесия. В случаях ацидоза избыточные ионы водорода мигрируют во внутрь клеток для буферирования. При этом, другой катион, а именно ион К+, покидает клетку и передвигается во внеклеточную жидкость. В случаях снижения уровня внеклеточного калия, ионы К+ перемещаются в клетки и заменяются ионами водорода. Таким образом, изменения уровней внеклеточного калия могут оказывать влияние на кислотно-основное состояние, и изменения кислотно-основного состояния влияют на уровни внеклеточного калия. Сдвиги калия, обыкновенно, более выражены при метаболическом ацидозе, чем при респираторном ацидозе. Следует отметить, что метаболический ацидоз, развивающийся в результате накопления неорганических кислот (например, HCl в случае диареи, фосфорной кислоты при почечной недостаточности), приводит к более значительному повышению калия во внеклеточном пространстве, чем ацидоз, вызываемый аккумуляцией органических кислот (например, лактата, кетокислот).
|
|
|
Органная регуляция КОС
1. Респираторные контрольные механизмы.
Изменение легочной вентиляции является одним из механизмов, обеспечивающих постоянство рН крови. Основная функция дыхательной системы, направленная на поддержание оптимального уровня напряжения кислорода и углекислого газа (рО2ирСО2), обеспечивается тремя процессами – вентиляцией, перфузией и диффузией газов в легких. При прохождении крови через легкие в эритроцитах и плазме крови осуществляются следующие реакции
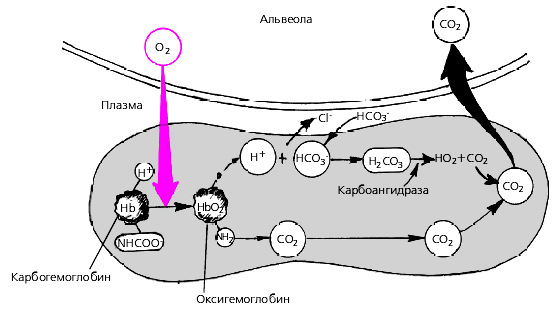
Рис.5. Химические реакции, протекающие в эритроцитах при газообмене в легких (По Р. Шмидту и Г. Ревсу и соавт., 1996).
При усиленном образовании и накоплении в организме ионов водорода бикарбонатная система связывает их с помощью бикарбоната натрия и переводит сильные кислоты в слабую угольную кислоту, которая диссоциирует на СО2 и Н2О. Углекислый газ выделяется с выдыхаемым воздухом. При удалении из крови углекислого газа в ней исчезает примерно эквивалентное число ионов водорода.
|
|
|

Избыток СО2 или избыток ионов Н+ в крови главным образом действует непосредственно на дыхательный центр в головном мозге, что приводит к изменению вентиляцию. Ионы Н+ с трудом проходят через гемато-энцефалический барьер, в то время как СО2 минует его с легкостью и в процессе взаимодействия с водой образует угольную кислоту, которая диссоциирует до ионов Н+ и НСО3ˉ. Ион Н+ стимулирует дыхательный центр, вызывая повышение или снижение вентиляции.
Респираторный контроль рН является быстро срабатывающим механизмом, осуществляющимся в течение нескольких минут (максимально – от 12 до 24 часов). Хотя респираторный ответ оперативен, он не позволяет полностью вернуть рН к нормальному уровню, и эффективен только на 50-75%. Это означает, что, если рН падает до 7,4 - 7,0, респираторная система способна приблизить рН к значениям около 7,2 – 7,3. Это быстрое действие, однако, предотвращает значительные изменения рН от момента их возникновения до вступления в ответ более медленно реагирующих почек.
Как отмечалось, СО2 легко пересекает гемато-энцефалический барьер, в отличие от замедленного перемещения НСО3ˉ ионов. Таким образом, уровни в крови рН и ионов НСО3ˉ снижаются более быстро, чем в ликворе. При метаболическом ацидозе, например, при котором имеет место первичное снижение НСО3ˉ ионов, максимальный респираторный ответ отсрочен до 12-24 часов. Когда метаболические кислотно-основные нарушения скорректированы быстро, гипервентиляция может сохраняться из-за отсрочки корректировки в церебро-спинальной жидкости.

Рис.6. Респираторная регуляция КОС.
Таким образом при самопроизвольном дыхании адекватные изменения вентиляции регулируются дыхательным центром, который реагирует на изменения содержания ионов водорода и углекислого газа. В условиях гиперкапнии и ацидоза развивается тахипноэ и гипервентиляция, СО2 выводится. При алкалозе возникает брадипноэ и гиповентиляция за счет снижения стимуляции дыхательного центра. Легочные механизмы обеспечивают временную компенсацию, так как при этом происходит смещение кривой диссоциации оксигемоглобина влево и уменьшается кислородная емкость артериальной крови.
Дата добавления: 2018-02-15; просмотров: 1638; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!