МОРФОГЕНЕЗ В КАЛЛУСНЫХ ТКАНЯХ
Существует несколько путей, по которым может идти развитие клетки после ее дедифференцировки. Первый путь — это вторичная регенерация целого растения, возможна дифференцировка на уровне клеток, тканей, органов. Второй путь — это утрата клеткой способности к вторичной дифференцировке и регенерации растения, стойкая дедифференцировка, приобретение способности расти на среде без гормонов, т.е. превращение в опухолевую. Такими свойствами часто характеризуются клетки старых пересадочных культур. Третий путь — это нормальный цикл развития каллусной клетки, заканчивающийся ее
29
|
|
старением и отмиранием. В этом случае клетка претерпевает вторичную дифференцировку и прекращает делиться (стационарная фаза
роста). Однако такая дифференцировка не ве-дет к морфогенезу, а закрепляет за ней свойства старой каллусной клетки.
Для сельскохозяйственной биотехнологии наибольший интерес представляет регенерация в культуре тканей из отдельной клетки целого растения. Иногда этот путь лежит через образование отдельных органов.
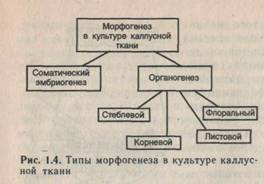
В культуре каллусных тканей морфогенезом называют возникновение организованных структур из неорганизованной массы клеток. Существует два основных типа морфогенеза (рис. 1.4). В культуре тканей он может проявляться в виде органогенеза (образования монополярной структуры, т.е. отдельных органов): корневого, стеблевого, реже флорального (цветочного) или листового, а также в виде соматического эмбриогенеза (образования биполярных зародышеподобных структур из соматических клеток). В случае органогенеза сначала регенерируют отдельные органы, а затем уже из них — целые растения, исключение составляет корневой органогенез.
|
|
|
В результате соматического эмбриогенеза в отличие от органогенеза сразу образуется зародыш, имеющий как меристему корня, так и меристему верхушечной почки, из которого в дальнейшем развивается целое растение.
Способность отдельной соматической клетки полностью реализовывать свою программу развития и давать начало целому растительному организму называют тотипотентностью растительной клетки. Любая растительная клетка обладает одинаковыми потенциальными возможностями, так как содержит весь набор генов и, следовательно, клетки сохраняют свойственную зиготе программу развития. Поэтому если мы получаем каллус из клеток лепестка цветка, или из клеток сердцевинной парен-хемы стебля, или из клеток любой ткани, то в принципе каждая такая клетка может регенерировать целое растение. Однако свойство тотипотентности не всегда реализуется, так как по-тенциальные возможности клеток разных типов проявляются
|
|
|
30
неодинаково. В некоторых из них гены в сильной степени репрессированы, в связи с чем проявление тотипотентности становится ограниченным.
Идея о тотипотентности растительной клетки была выдвинута Г. Хаберландтом еще в 1902 г., хотя и не получила тогда экспериментального подтверждения. Согласно определению Хаберландта, любая клетка растения может дать начало новому организму, и если этого не наблюдается, то только потому, что растительный организм подавляет потенции клетки к развитию. Изоляция клеток от растений способствует проявлению этих потенций.
Клеточную основу морфогенеза составляет цитодифферен-цировка. Регенерация растения начинается со вторичной дифференцировки клеток. При этом дедифференцированные клетки вновь приобретают структуру и функции специализированных.
Вторичная дифференцировка каллусных клеток не всегда заканчивается морфогенезом и регенерацией растения. Иногда она приводит только к образованию тканей (гистодифференцировка). Таким путем каллусная клетка может превращаться во флоэмные или ксилемные элементы. Другим примером вторичной дифференцировки может служить превращение дедифференцированной активно пролиферирующей клетки в старую неделящуюся каллусную клетку (стационарная фаза роста).
|
|
|
Из всех видов вторичной дифференцировки наибольший интерес представляет морфогенез, так как он позволяет получать целое растение из каллусной клетки.
В основе дифференцировки и морфогенеза лежит последовательное включение различных генов, т.е. дифференцировка клеток определяется дифференциальной активностью генов. Изменение активности структурных генов может быть связано с их дерепрессией, репрессией или амплификацией (умножением). Большую роль в этом процессе играют фитогормоны.
Морфогенезом в культуре каллусных тканей можно управлять. На способность изолированных растительных клеток к морфогенезу оказывают влияние как внутренние, так и внешние факторы. К внутренним факторам относятся: видовая принадлежность исходного растения, орган, из которого взят экс-плант, возраст экспланта. К внешним факторам прежде всего относятся: состав питательной среды, температура, свет (интенсивность и длина фотопериода). Наиболее мощным индуктором морфогенеза, который принято называть стимулом или сигналом морфогенеза, является изменение соотношения между цитокининами и ауксинами, входящими в состав питательных сред. При преобладании цитокининов над ауксинами часто на-
|
|
|
31
|
|
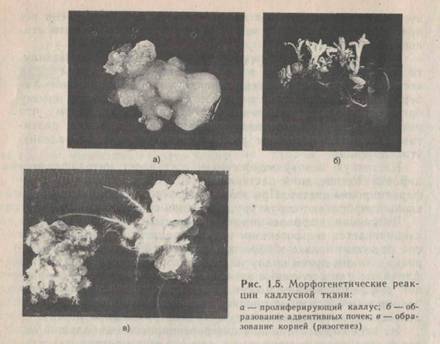
чинается стеблевой органогенез, а в случае преобладания ауксинов над цитокининами — корневой (рис. 1.5). Следует отметить, что из образующихся в культуре каллусной ткани корней почти никогда не регенерируется целое растение, а при стеблевом органогенезе сначала образуется побег, который затем (при пересадке на среду с преобладанием ауксинов) укореняется и дает начало целому растению.
Таким образом, различия в балансе экзогенных гормонов ауксинового и цитокининового типа определяет, с одной стороны, возможность перехода клетки в культуре к дедифференцировке и неорганизованной пролиферации, а с другой — индукцию вторичной дифференцировки того или иного типа морфогенеза, что было отмечено Ф. Скугом и Е. Миллером (1957). Следовательно, ауксины и цитокинины, вызывающие в зависимости от соотношения либо дедифференцировку и переход к каллусному росту, либо дифференцировку и морфогенез в культуре каллусных тканей, являются не только регуляторами роста, но и регуляторами дифференцировки.
32
Если органогенез можно индуцировать с помощью ауксинов или цитокининов, то соматический эмбриогенез фактически независим от экзогенных фитогормонов. Обычно эмбриогенные зоны возникают в каллусной ткани на той же питательной среде, которая использовалась для каллусообразования. Развитие соматических зародышей в каллусной ткани начинается тогда, когда устраняется дедифференцирующий фактор из питательной среды (2,4-Д или другие ауксины). Развивающийся зародыш не нуждается в экзогенных гормонах, так как сам обеспечивает себя ими.
Независимость соматического эмбриогенеза от гормонов является аргументом в пользу точки зрения, высказанной еще Хаберландтом, а позднее Стэвардом, что сам процесс изолирования клетки стимулирует реализацию ее тотипотентности, т.е. переход к морфогенезу. Таким образом, основными стимулами морфогенеза являются изменения соотношения гормонов в питательной среде, а также сам процесс изоляции растительной клетки от организма. Дополнительными стимулами морфогенеза в культуре каллусных тканей является присутствие в питательной среде нитрата серебра, нитрата аммония, некоторых аминокислот (пролин, тирозин, иногда серии), полиаминов (путресцин и спермидин). В ряде случаев стимулируют процесс морфогенеза маннит и сорбит. Ионы NO3-, оказывают влияние на развитие возникших в каллусной ткани организованных структур, а их индукцию стимулируют ионы NH+4. Гибберелловая кислота стимулирует рост зачатков стебля, а абсцизовая ускоряет дифференцировку органов соматических зародышей.
Интересно отметить, что некоторые из перечисленных веществ, например, нитрат серебра, продлевают регенерационную способность в старых пересадочных культурах.
Под влиянием того или иного стимула морфогенеза каллус-ная клетка должна стать детерминированной, однако не все клетки, а лишь одна из 400—1000 становится на путь регенерации. Следовательно, для перехода к морфогенезу недостаточно индуктора (стимула), а необходимо, чтобы клетка была готова к ответу на него. Способность воспринимать стимулы морфогенеза называют компетентностью клетки. Исследователи пришли к выводу, что компетентность клеток — событие случайное и поэтому столь редкое. В связи с этим напрашивается вопрос о судьбе тех каллусных клеток, которые в силу некомпетентности не способны воспринять стимулы морфогенеза и детерминироваться. В пересадочной культуре эти клетки продолжают делиться и скорее всего становятся на путь перехода к гормононезависимости. Однако не все каллусные ткани со временем
2-17733
|
|
завершают развитие возникновением гормононезависимости. Многие из них в силу генетических особенностей продолжают использовать экзогенные гормоны, но полностью утрачивают способность к регенерации. Такие ткани занимают промежуточное положение между «привыкшими» и свежими каллусными тканями.
Морфогенез в каллусной ткани начинается с того, что под влиянием соответствующих условий детерминированная клетка обособляется от окружающих ее каллусных клеток, образуя утолщенную клеточную стенку. Это явление было обнаружено в 1972 г. Данилиной при изучении соматического эмбриогенеза в культуре ткани моркови.
Клетка-инициаль при соматическом эмбриогенезе дает начало зиготе, а при органогенезе — меристематическому очагу. От недетерминированных каллусных клеток инициальная отличается более крупным ядром и меньшими размерами вакуолей. Ядро обычно занимает центральное положение. В инициальных клетках содержатся большие количества запасных веществ: крахмала, иногда—липидов.
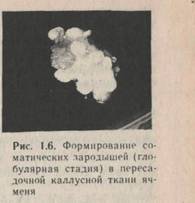
Некоторое время инициальные клетки находятся в лаг-фазе, что необходимо для их перестройки и подготовки к последующим быстрым делениям. Затем эти клетки делятся по ти-пу дробления, образуя сферическую массу мелких изодиаметрических клеток. В случае органогенеза эту массу клеток называют меристематическим очагом, а в случае соматического эмбриогенеза — глобулярным проэмбрио. В дальнейшем в меристематическом очаге дифференцируются зачатки стебля, корня, листа или цветочной почки и соответственно происходит стеблевой, корневой, листовой или флоральный органогенез. В глобулярном проэмбрио развивается биполярная эмбриоидная структура. Можно выделить несколько последовательных стадий формирования соматических эмбриоидов из каллусной клетки: глобулярную, сердечка, торпедовидную, соматического зародыша. Меристематические очаги или проэмбрио могут возникать на периферии каллусной ткани или быть погруженными в нее. Обычно не наблюдается определенной закономерности в их локализации (рис. 1.6). Исключение составляет стеблевой органогенез в каллусной ткани, получен-ной из сердцевинной паренхимы табака сорта Висконсин-38. Меристематические очаги здесь всегда локализуются только в нижней части каллусной массы.
При переходе каллусных клеток к морфогенезу происходит существенное изменение их метаболизма. Морфогенезу предшествует появление в клетках белков-антигенов. Работами
34
Р.Г. Бутенко, Н.И. Володарского и Н.А. Моисеевой показано, что морфогенез в культуре каллусных тканей табака характеризуется включением и выключением синтеза определенных белков-маркеров. В меристемах обнаружено два белка-антигена, которые являются маркерами этих клеток. Одновременно показано, что индуцированная детерминация клеток каллусной ткани сопряжена с появлением в ней антигена-маркера клеток меристемы стебля.
Белок (гликопротеид), выделенный из эмбриогенных культур, можно рассматривать как конденционирующий фактор. При частых пересадках на свежую питательную среду, где гликопротеид накапливаться не может, эмбриогенез не идет. Если белок, появляющийся в клетках при переходе к соматическому эмбриогенезу, выделить и ввести в длинные (неэмбриогенные) каллусные клетки, у которых гены морфогенеза не работают или потеряны, то в них индуцируется переход к морфогенезу.
Работы по поиску новых маркеров морфогенеза продолжаются. Клетки меристематических очагов и клетки, дающие начало эмбриоидным структурам, отличаются от каллусных интенсивным синтезом РНК и ДНК, что связано с особенностями их белкового обмена. Изменения в белковом обмене сходны с теми, которые протекают при дедифференцировке клетки, но итоги у них различны. По мнению Р. Г. Бутенко, специфика реакции определяется не общим усилением синтеза макромолекул, что необходимо для усиленной пролиферации, а теми уникальными синтезами, которые идут на этом общем фоне и обусловливают появление белков регуляторного типа.
Переход к морфогенезу в культуре каллусных тканей сопро-вождается значительными изменениями дыхательного метаболизма. В целом дыхание (по С02) усиливается, но изменяется его характер в направлении интенсификации пентозофосфатного пути. Возрастает активность дыхательных ферментов.
Вслед за биохимической наступает структурная реорганизация клетки. Биохимическая дифференцировка клетки всегда предшествует структурной. В клетках, вступивших на путь морфогенеза, возрастает число рибосом, митохондрий, меняется их внутренняя структура.
Процессы морфогенеза в каллусных клетках протекают асинхронно и продолжительно. Одновременно в каллусной тка-
35
ни могут иметься как полностью сформированные структуры, так и клетки, только что вступившие на этот путь.
Повышенная синтетическая активность клеток меристематического очага и глобулярного проэмбрио делает их аттрагирующим центром, в который устремляются питательные вещества. Окружающие каллусные клетки при этом часто разрушаются и образующиеся эмбриоиды легко выпадают из массы каллусных клеток.
Каллусные клетки не связаны между собой плазмодесмами или последние сильно редуцированы. При появлении зароды-шеподобных структур или меристематических очагов между клетками снова восстанавливается связь с помощью плазмодесм.
Все изменения, происходящие при морфогенезе и заканчивающиеся регенерацией из каллусой клетки растения, управляются (контролируются) специальными генами. В настоящее время одни ученые считают, что признак морфогенеза полигенен и контролируется несколькими хромосомами, другие при-шли к заключению, что этот признак определяется двумя ядерными генами. Тот факт, что морфогенетическая активность каллусных клеток имеет генетическую природу, объясняет, почему не удается в ряде случаев получить регенерацию из каллусной ткани тех или иных генотипов. Регенерационную способность может увеличить скрещивание генотипов, морфогенетически активных invitro.
Дата добавления: 2021-01-20; просмотров: 251; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!