Реакции на аскорбиновую кислоту (витамин С)
Принцип
Аскорбат обладает восстанавливающими свойствами и может восстанавливать 2,6‑дихлорфенолиндофенол, метиленовый синий, калия гексацианоферрат К3Fе(СN)6, окисляясь при этом до дегидроаскорбата. 2,6‑Дихлорфенол-индофенол и метиленовый синий восстанавливаются до бесцветных лейкосоединений, а К3Fе(СN)6 восстанавливается до К4Fе(СN)6, дающего с ионами трехвалентного железа соль Fe4[Fе(СN)6] сине-зеленого цвета.
Проведение анализа
1 реакция
В пробирку вносят 10 капель раствора 2,6‑дихлорфенолиндофенола, 1‑2 капли 10 % раствора HСl и по каплям 1 % раствор аскорбиновой кислоты до обесцвечивания 2,6‑дихлорфенолиндофенола.
2 реакция
В две пробирки вносят по капле метиленового синего. В первую прибавляют 5 капель раствора аскорбиновой кислоты, во вторую 5 капель дистиллированной воды и ставят в водяную баню (+40°С). Через некоторое время в пробирке с витамином жидкость обесцвечивается.
3 реакция
К 10 каплям аскорбиновой кислоты прибавляют 10 капель раствора калия гексацианоферрата К3Fе(СN)6 и 5 капель раствора FeCl3. Наблюдают образование сине-зеленого окрашивания (берлинская лазурь).
Практическое значение
Качественные реакции на витамины позволяют установить подлинность (достоверность) витаминных лекарственных препаратов, также реакции можно использовать для обнаружения и количественного определения витаминов в пищевых объектах и лекарственных растениях.
|
|
|
Оформление работы
Указывают принципы используемых в работе методов, регистрируют полученные результаты в виде приведённой ниже таблицы, делают вывод о наличии или отсутствии каждого из изучаемых водорастворимых витаминов в исследуемом материале.
| Название исследуемого витамина | Окраска | Вывод | |||
| В1 | Тиамин | ||||
| В2 | Рибофлавин | реакция восстановления |
| ||
| реакция окисления | |||||
| ФМН (рибофлавиннуклеотид) | |||||
| ФАД (флавинат) | |||||
| В5 | Никотинамид | ||||
| В6 | Пиридоксин | ||||
| В12 | Кобаламин | ||||
| С | Дигидро-аскорбат | реакция с 2,6‑ДХФИФ |
| ||
| реакция с метиленовым синим | |||||
| реакция с К3Fе(СN)6 | |||||
Вопросы для самоконтроля
1) Строение жирорастворимых витаминов А, Е, D, их активные формы.
2) Строение водорастворимых витаминов с указанием активного центра (В1, В2, В6, РР, С, Н), их активные формы (ТДФ, ФМН, ФАД, ПФ, НАД+, НАДФ+).
3) Представление о химическом строении водорастворимых витаминов В3, В12, В9(ВС), их активных форм, жирорастворимых витаминов К, F и коэнзима Q.
4) Классификация и номенклатура витаминов, единицы активности. Про- и антивитамины, витаминоподобные вещества.
|
|
|
5) Витамины в питании. Факторы, влияющие на обеспеченность организма витаминами. Депонирование и выведение витаминов.
6) Характеристика отдельных жиро‑ и водорастворимых витаминов, витаминоподобных веществ, их биологическая роль, клиническая картина авитаминозов (гипервитаминозов, если есть), суточная потребность, источники.
7) Витамины и антивитамины как лекарственные вещества.
Тестовые задания
Выбрать один правильный ответ.
1. Основная функция водорастворимых витаминов – это
1) защита транспорта субстратов в клетки
2) защита биологических мембран
3) предшественники коферментов
4) предшественники углеводов
5) предшественники гормонов
2. Пантотеновая кислота является составной частью кофермента
1) коэнзим А
2) тетрагидрофолиевая кислота
3) тиаминпирофосфат
4) флавинмононуклеотид
5) коэнзим Q
3. При длительном приеме антибиотиков, сульфаниламидов развивается гиповитаминоз В6, обусловленный
1) подавлением микрофлоры кишечника
2) связыванием лекарства с витамином
3) действием лекарства на синтез коферментной формы
4) ингибированием пиридоксин-зависимых ферментов
5) блокированием трансмембранного переноса витамина
|
|
|
4. При дефиците аскорбиновой кислоты развивается цинга вследствие
1) окисления сульфгидрильных групп ферментов
2) нарушения синтеза коллагена
3) нарушения синтеза альбумина
4) окисления липидных мембран клеток соединительной ткани
5) нарушения транспорта кальция
5. В состав витамина F входят жирные кислоты
1) олеиновая, линолевая и линоленовая
2) олеиновая, линолевая и стеариновая
3) арахидоновая, линолевая и линоленовая
4) олеиновая, линолевая и арахидоновая
5) олеиновая, линоленовая и арахидоновая
6. Основная роль витамина К состоит в том, что он
1) является антиоксидантом
2) увеличивает образование тромбоцитов
3) участвует в синтезе факторов свертывания крови
4) участвует в реакциях протеолиза
5) участвует в реакциях минерализации
7. Фермент в отличие от небелкового катализатора
1) снижает энергию активации
2) не расходуется в результате реакции
3) не претерпевает необратимых изменений
4) обладает специфичностью
5) требуется в малом количестве
8. Перенос групп внутри молекулы катализируют ферменты
1) изомеразы
2) трансферазы
3) лиазы
4) гидролазы
5) лигазы
9. Субстратная специфичность ферментов обусловлена
1) набором определенных функциональных групп в активном центре
|
|
|
2) химическим соответствием активного центра субстрата
3) наличием кофермента
4) пространственным соответствием активного центра субстрата
5) комплементарностью активного центра субстрату
10. Действие конкурентного ингибитора можно снять, если
1) увеличить концентрацию ингибитора
2) увеличить концентрацию субстрата
3) снизить концентрацию фермента
4) изменить условия реакции – рН и температуру
5) снизить коллоидоустойчивость фермента
Ситуационные задачи
1. Обнаружено, что если аллостерический фермент аспартат:карбамоил-трансфераза (молекула которого состоит из 12 протомеров) выдержать в течение 4 минут при 60°С, то он теряет чувствительность к аллостерическому ингибитору (ЦТФ). При этом ферментативная активность сохраняется. Схожие свойства проявляют и другие аллостерические ферменты. Предложить возможные механизмы подобного нарушения.
2. Липаза – фермент жировой ткани, обеспечивающий расщепление нейтральных жиров, может находиться в двух формах с различной активностью: в виде простого белка и в виде фосфопротеина. Объяснить, почему переход одной формы в другую сопровождается изменением активности. Предположить, в каком состоянии липаза является активной, если известно, что выделяющийся при физической нагрузке гормон адреналин запускает каскад реакций, ведущих к фосфорилированию внутриклеточных белков.
 Строение сульфаниламидов Строение сульфаниламидов
|
3. В качестве антибактериальных средств широкого спектра действия первыми стали использовать сульфаниламиды, содержащие структуру, схожую с парааминобензойной кислотой. Указать, на чем основано использование сульфаниламидов и что ещё обычно рекомендуют при их применении
4. Бактерии Lactobacillus casei растут на простой культуральной среде, содержащей витамины рибофлавин и пиридоксин и 4 аминокислоты. Если в культуральную среду добавить полный набор аминокислот и рибофлавин, то количество пиридоксина, необходимого для оптимального роста бактерий, сократится на 90 %. Пояснить, почему это происходит.
ТЕМА
"Белки. Нуклеиновые кислоты. Ферменты. Витамины"
(контрольное занятие по разделам 1 и 2)
Вопросы для самоподготовки
Белки . Нуклеиновые кислоты
1. Понятие о белках. Биологическая роль белков.
2. Аминокислотный состав белков. Строение 20 природных аминокислот, классификация по строению радикала. Функциональные группы аминокислот, их роль. Заменимые и незаменимые аминокислоты.
3. Индивидуальная роль аминокислот вне состава белков. Аминокислоты ‑ лекарственные препараты.
4. Физико-химические свойства белков, очистка и разделение, виды осаждения. Высаливание, его механизм, практическое значение. Необратимое осаждение белков, агенты и механизмы, использование в биохимии и медицине.
5. Уровни структурной организации белковой молекулы. Типы связей, стабилизирующих структуру белковой молекулы. Де- и ренатурация белков.
6. Составить и назвать пептид, определить суммарный заряд, растворимость, зону pH с изоэлектрической точкой, движение в электрическом поле.
7. Классификация белков по функциям (примеры).
8. Классификация белков по строению. Белки простые и сложные.
9. Представление о структуре и биологической роли фосфо‑, глико‑, липо‑, металло‑, хромопротеинов (гемо‑, флаво‑ и ретиналь‑протеинов).
10.Нуклеопротеины, структура белковой и небелковой части. Особенности строения гистонов и протаминов, роль в стабилизации нуклеиновых кислот.
11.Структурные формулы азотистых оснований. Нуклеозиды. Классификация, структура нуклеотидов, представленных в нуклеиновых кислотах.
12.Понятие о нуклеотидах, отличных от нуклеотидов нуклеиновых кислот. Роль отдельных моно‑ и динуклеотидов (участие в активации молекул, детоксикации, катализе, регуляции, энергетике). Строение, роль циклонуклеотидов.
13.Биологические и метаболические взаимосвязи аминокислот и нуклеиновых кислот. Информация о белке. Аминокислоты ‑ источники синтеза и конечные продукты распада нуклеотидов. Мочевая кислота ‑ представитель пуринов.
14.Виды нуклеиновых кислот, локализация в клетке, биологическая роль, сходство и различия их первичной и вторичной структур.
15.Типы РНК, их локализация, функции, особенности строения, уровни организации, стабилизирующие связи.
16.ДНК: физико-химические свойства, состав, уровни организации, стабилизирующие связи, биологическая роль. ДНК про-, эукариот, ядра, митохондрий.
Ферменты. Витамины
1. Понятие о ферментах. История изучения энзимологии.
2. Химическая природа ферментов. Сходство и различия между ферментами и неферментными катализаторами.
3. Проферменты, изоферменты, множественные формы фермента.
4. Структурно-функциональная организация ферментов: уровень структуры, простые и сложные ферменты, холофермент, роль апофермента и кофактора (кофермента, простетической группы) в катализе, активный и аллостерический центры.
5. Кинетика ферментативных процессов. Механизмы катализа: ковалентный, кислотно-основной и др.
6. Свойства ферментов: специфичность, влияние температуры, pH среды, количества субстрата; активация и ингибирование, типы и обратимость ингибирования.
7. Регуляция активности ферментов: аллостерические механизмы, фосфорилирование и дефосфорилирование, частичный протеолиз, белок-белковые взаимодействия, компартментализация и др.
8. Принципы современной номенклатуры и классификации ферментов. Характеристика ферментов по схеме: класс, основные подклассы, строение, коферменты, катализируемая реакция, биологическая роль.
9. Принципы количественного определения активности ферментов. Единицы активности ферментов.
10. Использование ферментов в медицине: энзимодиагностика, энзимопатология, энзимотерапия (ферменты и ингибиторы ферментов). Значение определения изоферментов в клинической практике.
11.Признаки витаминов и витаминоподобных веществ. Ученые ‑ основоположники витаминологии. Классификация и номенклатура витаминов.
12.Гипо- и авитаминоз экзо‑ и эндогенный. Гипервитаминоз.
13.Провитамины и антивитамины (примеры).
14.Строение витаминов А, D3, Е, С, Н, В1, В2, В6, РР, активные центры, коферментная роль биологически активных форм (ФМН/ФАД, НАД+/НАДФ+, ТДФ, ПФ и др.). Представления о строении, роли витаминов К, F, В3, В12, ВС.
15.Характеристика витаминов по плану: 1) химическая структура, активная форма; 2) источники, образование из провитамина; 3) суточная потребность; 4) биологическая роль; 5) недостаточность, гипервитаминоз (если есть).
16.Витамины ‑ лекарственные препараты.
Практическая часть
1. Исследование белка и свободных аминокислот в биологическом материале.
2. Исследование высаливания и денатурации белков.
3. Выделение и анализ компонентов фосфо-, глико-, нуклеопротеинов.
4. Обнаружение небелкового компонента гемопротеинов, мочевой кислоты.
5. Зависимость скорости ферментативной реакции от температуры.
6. Влияние активаторов и ингибиторов на активность амилазы.
7. Оценка специфичности действия уреазы.
8. Определение активности фермента.
9. Оценка работы ферментов разных классов.
Раздел 3.
Введение в обмен веществ и энергии.
Биологическое окисление
ТЕМА 3.1.
Введение в обмен веществ и энергии. Общие пути катаболизма и окислительное фосфорилирование (семинар)
Актуальность
Биологическое окисление протекает во всех живых клетках организма в виде совокупных окислительных реакций. Основной функцией этого процесса является обеспечение организма энергией для процессов жизнедеятельности. Основной формой запасания энергии, доступной для использования, является аденозинтрифосфорная кислота (АТФ). Синонимом биологического окисления является тканевое дыхание. Процесс биологического окисления протекает многостадийно с участием промежуточных ферментативных реакций. Происходит многократная передача протонов и электронов или только электронов от донора к акцептору. Конечным акцептором электронов и протонов служит ½О2. Конечными продуктами тканевого дыхания являются вода окисления и диоксид углерода (Н2О и СО2). Процесс фосфорилирования, сопряженный с тканевым дыханием, называется окислительным фосфорилированием. Чрезвычайно важной функцией цепи дыхательных ферментов митохондрий наряду с передачей электронов является аккумуляция части освобождающейся энергии в фосфатных связях макроэргических соединений. В организме существуют альтернативные биологическому окислению пути использования кислорода.
Цель
1) Изучить роль кислорода в организме, пути его использования.
2) Ознакомиться с процессами биологического окисления и конечными продуктами тканевого дыхания.
3) Изучить механизмы окислительных реакций в клетке и строение основных коферментов тканевого дыхания.
4) Изучить механизмы окислительного фосфорилирования и пути образования макроэргов с оценкой количества образующейся АТФ в ходе тканевого дыхания, сопряженного с окислительным фосфорилированием.
Вопросы для самоподготовки
1) Понятие об обмене веществ. Пластическая и энергетическая роль метаболизма.
2) Взаимосвязь катаболизма и анаболизма, энергетика и субстраты процессов.
3) Этапы катаболизма. Стадии катаболических превращений питательных веществ в организме, связанные с высвобождением свободной энергии.
4) Формы трансформации свободной энергии — образование восстановленных эквивалентов (НАДФН×Н) и синтез "макроэргических" соединений (АТФ, 1,3‑дифосфоглицерат, креатинфосфат, ацил~SКоА и др.).
5) Циклы АТФ–АДФ, НАДФН×Н–НАДФ. Свойства молекул АТФ и НАДФН. АТФ – универсальный источник и переносчик химической энергии в клетке.
6) Пути участия кислорода в метаболизме.
7) Понятие о тканевом дыхании (биологическом окислении) и локализации ферментов тканевого дыхания на внутренней мембране митохондрий.
8) Совокупность процессов тканевого дыхания. Субстраты окисления.
9) Строение мультиферментного пируватдегидрогеназного комплекса, ферменты, роль коферментов и работа их активных центров. Механизм окислительного декарбоксилирования пирувата.
10) Ацетил~КоА ‑ ключевая фигура метаболизма.
11) Цикл трикарбоновых кислот (ЦТК, цикл Кребса) ‑ универсальный механизм окисления. Схема взаимосвязей ЦТК с распадом углеводов, липидов, белков.
12) Химические реакции цикла Кребса. Дегидрогеназы ЦТК и их коферменты. Механизм субстратного фосфорилирования в ЦТК. Биологическая роль цикла Кребса.
13) Роль кислорода как конечного акцептора протонов и электронов восстановленных субстратов биологического окисления.
14) Строение внутренней мембраны митохондрий и компонентов дыхательной цепи. Последовательность компонентов цепи переноса электронов. Молекулярная организация ферментных ансамблей, роль коферментов. ФМН, FeS‑белки, коэнзим Q, гемовые группы цитохромов.
15) Энергетическая лестница компонентов дыхательной цепи. Изменение свободной энергии в процессе переноса электронов, энергетические перепады и образование АТФ.
16) Трансмембранный перенос Н+, механизм окислительного фосфорилирования. Представление о протонной АТФ‑синтетазе, транслоказах АТФ и АДФ.
17) Коэффициент Р/О и метод его вычисления.
18) Сходство и отличия субстратного и окислительного фосфорилирования.
19) Возможности регуляции сопряжения окисления и фосфорилирования, физиологическое значение.
20) Причины разобщения дыхания и фосфорилирования, примеры действия разобщителей. Блокаторы тканевого дыхания, действие и примеры влияния на организм.
21) Неполное восстановление кислорода до воды, образование активных форм кислорода. Значение НАДФН как метаболической восстановительной энергии. Влияние на процессы метаболизма.
22) Понятие о микросомальном окислении, свободно-радикальном перекисном окислении макромолекул клетки и системе антиоксидантной защиты. Суть альтернативности процессов биологического окисления и перекисного окисления.
Самостоятельная работа
Составить схему энергетического обмена. На схеме представить химизм и взаимосвязи общих путей катаболизма: пируватдегидрогеназный комплекс, цикл трикарбоновых кислот, энергетическую цепь внутренней мембраны митохондрий. Приложить к схеме суммарные реакции показанных путей обмена.
Тестовые задания
Выбрать один правильный ответ.
1. Скорость пируватдегидрогеназной реакции ингибируют
1) АТФ, кальций, НАД+
2) кальций, ацетил-КоА, НАД+
3) АДФ, ФАДН2, НАДН
4) ацетил-КоА, НАДН, АТФ
2. В цикле трикарбоновых кислот молекула ФАДН2 образуется при работе
1) малатдегидрогеназы
2) изоцитратдегидрогеназы
3) сукцинатдегидрогеназы
4) a-кетоглутаратдегидрогеназы
3. Скорость цикла трикарбоновых кислот определяется концентрацией
1) a-кетоглутарата
2) оксалоацетата
3) янтарной кислоты
4) цитрата
5) яблочной кислоты
4. Движущей силой переноса электронов по цепи дыхательных ферментов является
1) энергия распада АТФ
2) перекачивание протонов водорода через мембрану
3) работа железосерных центров
4) различная электроотрицательность переносчиков
5. В дыхательном контроле проявляется зависимость скорости переноса электронов по дыхательной цепи от
1) соотношения концентрации АДФ и АТФ
2) концентрации НАДН
3) величины потребляемого кислорода
4) активности АТФ-синтетазы
6. Увеличение электрохимического градиента приведет
1) к увеличению скорости перекачивания протонов
2) к ускорению синтеза АТФ
3) к повышению скорости переноса электронов
4) к повышенному выделению СО2 и Н2О
7. Энергия, высвобождаемая при переносе электронов по цепи дыхательных ферментов, используется на
1) перекачивание ионов Н+ через мембрану
2) окисление железосерных центров
3) образование молекул воды
4) синтез АТФ
8. Создание протонного градиента на митохондриальной мембране обусловлено
1) распадом АТФ
2) окислением НАДН
3) движением электронов
4) выкачиванием ионов Н+ в обмен на Na+
5) изменением редокс-состояния фосфолипидов
9. Внедрение разобщителя в митохондриальную мембрану приведет
1) к снижению окисления НАДН
2) к активации синтеза АТФ
3) к снижению переноса электронов по дыхательной цепи
4) к увеличению протонного градиента
10. Образующиеся в цикле трикарбоновых кислот восстановленные эквиваленты используются
1) в цепи дыхательных ферментов
2) в реакциях синтеза глюкозы, жирных кислот
3) для работы АТФ-синтетазы
4) для синтеза ацетил-КоА
Ситуационные задачи
Ответы подробно и мотивированно объяснить.
1. Прием внутрь разобщающих агентов вызывает обильное потоотделение и повышение температуры тела. Дайте этому объяснение на молекулярном уровне. Как изменяется соотношение Р/О в присутствии разобщающих агентов? Можно ли использовать разобщители для борьбы с ожирением?
2. Особая жировая ткань – бурый жир – имеется у новорожденных, а также у некоторых животных, впадающих в зимнюю спячку или приспособленных к обитанию в холодных регионах планеты. В митохондриях бурого жира выход АТФ на 1 атом поглощенного кислорода составляет менее 1 молекулы, в то время как других тканях 2-3 молекулы. Какая физиологическая функция может определяться таким низким отношением Р/О в буром жире новорожденных? Каковы возможные механизмы, способные пределять столь низкое отношение Р/О, которое характерно для митохондрий бурого жира?
Раздел 4.
Строение, функции, обмен углеводов
ТЕМА 4.1.
Строение, внешний обмен, депонирование углеводов
Актуальность
В организме человека и животных углеводы играют важную роль и выполняют разнообразные функции: служат источником энергии, обеспечивая до 67 % суточного энергопотребления организма; являются пластическим материалом клеток и межклеточного вещества; используются в качестве исходных продуктов для синтеза липидов, белков и нуклеиновых кислот; участвуют в поддержании иммунитета (углеводные компоненты иммуноглобулинов); участвуют в рецепции и клеточных контактах (углеводные компоненты клеточной мембраны). Основной источник углеводов организма ‑ различные пищевые продукты, главным образом, растительного происхождения. Суточная норма потребления углеводов 450‑500 г. Поступающие в организм углеводы перевариваются в желудочно-кишечном тракте и всасываются в кровь в виде моносахаридов. В тканях часть глюкозы откладывается в виде гликогена. Особая роль в рационе питания принадлежит неперевариваемым и неусваиваемым полисахаридам, в первую очередь, целлюлозе.
К заболеваниям, связанным с патологией углеводного обмена, относят сахарный диабет, гликогенозы, мукополисахаридозы, галактоземию, фруктоземию, наследственную непереносимость ряда углеводов.
Цель
1. Изучить переваривание углеводов в пищеварительном тракте.
2. Изучить биосинтез и мобилизацию гликогена как энергетического резерва организма.
3. Практически оценить влияние пищеварительных соков на углеводы пищи.
Вопросы для самоподготовки
1. Структура основных представителей моно‑, ди‑, полисахаридов (рибоза, глюкоза, фруктоза, галактоза, мальтоза, лактоза, сахароза, крахмал, гликоген, целлюлоза, декстраны, леваны). Их происхождение, биологическая роль. Сходство и отличие структур крахмала, гликогена, целлюлозы.
2. Углеводы, поступающие в организм с пищей. Суточная потребность в углеводах.
3. Ферменты пищеварительных соков, участвующие в переваривании углеводов: класс, подкласс, локализация синтеза. Комплексы пристеночного пищеварения.
4. Реакции, катализируемые a‑амилазой, конечной декстриназой, мальтазой, лактазой, сахаразой. Конечные продукты реакций.
5. Причины неперевариваемости целлюлозы в желудочно-кишечном тракте человека. Роль целлюлозы в рационе питания.
6. Реакции синтеза и распада гликогена. Физиологическое значение и соотношение процессов в зависимости от ритма питания и режима работы мышц. Особенности обмена гликогена в печени и мышцах.
7. Аденилатциклазный механизм регуляции активности ферментов обмена гликогена. Влияние адреналина, глюкагона и инсулина на концентрацию цАМФ в клетке.
8. Участие ионов Са2+ в регуляции активности ферментов обмена гликогена.
9. Биохимические основы наследственных нарушений переваривания углеводов и обмена гликогена (лактозная интолерантность, непереносимость сахарозы, гликогенозы и агликогенозы).
10. Особенности утилизации простых сахаров микрофлорой полости рта. Понятие «метаболический взрыв», направления синтеза полимерных углеводов.
Самостоятельная работа
Дополнить схему энергетического обмена (раздел 3) в ходе изучения раздела 4 до общей схемы углеводно-энергетического обмена. По теме 4.1 на схеме представить химизм и взаимосвязи гликогеногенеза и гликогенолиза, их выход на общие пути катаболизма (пируватдегидрогеназный комплекс и цикл трикарбоновых кислот с энергетической цепью внутренней мембраны митохондрий). Приложить к схеме суммарные реакции показанных путей обмена.
Лабораторная работа 1.
Исследование влияния амилазы
на крахмал и целлюлозу
Крахмал является гомополисахаридом, состоящим из a‑амилозы, амилопектина. В a‑амилозе остатки глюкозы связаны между собой a‑(1,4)‑гликозидными связями, в амилопектине ‑ a‑(1,4)‑гликозидными и a‑(1,6)‑гликозидными связями. Целлюлоза является гомополисахаридом, в котором остатки глюкозы связаны между собой b‑(1,4)‑гликозидными связями. Фермент a‑амилаза гидролизует только a‑(1,4)‑гликозидныесвязи.
Принцип
Ферменты, содержащиеся в биологических жидкостях, расщепляют полисахариды. Продукты переваривания полисахаридов определяют с помощью реакции Троммера, выявляющей у высвобождаемых при гидролизе сахаров (глюкозы и мальтозы) способность к восстанавлению Cu(OH)2 голубого цвета в CuOH желтого цвета и CuO красного цвета при нагревании в щелочной среде.
Материал для исследования
Слюна, разведение (1:5).
Реактивы
1) Желудочный сок, 2) 5 % раствор панкреатина, 3) 1 % раствор крахмала, 4) 1 % водная суспензия целлюлозы, 5) 1 % раствор CuSO4; 6) 10 % раствор NaOH.
Проведение анализа
Готовят пробы соответственно таблице.
| N пробы | Крахмал, мл | Суспензия целлюлозы, мл | Слюна, мл | Желудочный сок, мл | Панкреатин, мл |
| 1 2 3 4 5 6 7 8 | 1,0 1,0 1,0 1,0 ‑ ‑ ‑ ‑ | ‑ ‑ ‑ ‑ 1,0 1,0 1,0 1,0 | 1,0 ‑ 1,0 ‑ 1,0 ‑ 1,0 ‑ | ‑ 1,0 1,0 ‑ ‑ 1,0 1,0 ‑ | ‑ ‑ ‑ 2,0 ‑ ‑ ‑ 2,0 |
Пробирки инкубируют 30 мин при 37°С в термостате или водяной бане. После инкубации проводят анализ на присутствие продуктов расщепления (глюкозы и мальтозы) полисахарида с помощью реакции Троммера. В каждую из 8 пробирок добавляют по 1 мл раствора NaOH и по 5 капель раствора CuSO4. Все пробирки ставят в кипящую водяную баню, кипятят 1 мин. Появление красного осадка меди оксида (I) указывает на положительную реакцию Троммера в присутствии глюкозы, мальтозы.
Оформление работы
Указывают принцип метода, итоги работы оформляют в виде таблицы:
| № проб | Субстрат | Источник фермента | Фермент | Результат |
Делают вывод об особенностях переваривания крахмала и целлюлозы в пищеварительном тракте, объясняют особенности переваривания углеводов и указывают причины различий.
Лабораторная работа 2.
Исследование влияния амилазы на крахмал и сахарозу
Реактивы
1) 1 % раствор крахмала, 2) 1 % раствор сахарозы, 3) рабочий реактив Фелинга (ex tempore): смешать по 10 капель реактивы Фелинг I, Фелинг II.
Материал для исследования
Слюна, разведение 1:10 (источник a‑амилазы).
Принцип
Метод основан на сравнительном изучении способности фермента амилазы к гидролизу разных углеводных субстратов: полисахарида крахмала и дисахарида сахарозы. Действие фермента на субстрат выявляют при помощи качественной реакции на свободную альдегидную группу углеводов (реакция Фелинга). Крахмал и сахароза не имеют свободной альдегидной группы, поэтому не дают положительной реакции с реактивом Фелинга. Реакция может быть положительной (появление красно-оранжевой окраски) только в случае расщепления этих субстратов до мальтозы и глюкозы, которые имеют свободную альдегидную группу и обладают восстанавливающими свойствами.
Проведение реакции
Берут две пробирки. В одну добавляют 10 капель крахмала, в другую — 10 капель раствора сахарозы. Добавляют в каждую пробирку по 5 капель разбавленной слюны, перемешивают и ставят на 10 мин в водяную баню или термостат (37°С). Проделывают с содержимым обеих пробирок реакцию Фелинга: к 5 каплям исследуемого раствора добавляют 3 капли рабочего реактива Фелинга (приготовленного самостоятельно в отдельной пробирке), пробы нагревают до кипения на водяной бане и кипятят 1 мин. Сравнивают окраску в пробирках.
Оформление работы
Указывают принцип метода, регистрируют результаты в таблице:
| № проб | Субстрат | Источник фермента | Фермент | Результат |
| 1 | ||||
| 2 |
Делают вывод о специфичности действия a‑амилазы на субстрат, возможности действия на сахарозу и о локализации фермента в пищеварительном тракте.
Вопросы для самоконтроля
1. Классификация углеводов по структуре и функциям. Представители.
2. Переваривание и всасывание углеводов пищи. Ферменты полостного пищеварения, пристеночные комплексы. Наследственная непереносимость углеводов, дефектные ферменты. Роль целлюлозы в пищеварении.
3. Реакции инверсии пищевых моносахаров в энтероците. Универсальность молекулы глюкозы.
4. Транспорт глюкозы через мембраны, условия поступления в клетку, регуляция, способ депонирования.
5. Реакции синтеза и распада гликогена. Гормональная регуляция обмена гликогена. Особенности обмена гликогена в печени и мышцах. Гликогенозы.
6. Утилизация простых сахаров микрофлорой полости рта в процессе «метаболического взрыва.
ТЕМА 4.2.
Анаэробный и аэробный обмен углеводов
Актуальность:
Гликолиз ‑ центральный путь катаболизма глюкозы, может поставлять углеродный скелет для процессов биосинтеза. В анаэробных условиях гликолиз ‑ единственный процесс в организме, поставляющий энергию. Именно благодаря гликолизу организм может осуществлять ряд физиологических функций в условиях гипоксии, платой за это становится накопление лактата. У некоторых анаэробных организмов, таких как дрожжи, а также в некоторых клетках высших организмов (например, эритроцит) анаэробное превращение углеводов является единственным источником АТФ. В аэробных условиях при отсутствии НАДН гликолиз не может быть завершён и становится начальной стадией расщепления энергетических субстратов (до промежуточного продукта – пирувата), которую продолжают реакции аэробного окисления до СО2, Н2О и АТФ.
Аэробный распад глюкозы ‑ основной путь ее катаболизма у аэробных организмов, который поставляет гораздо больше энергии, чем анаэробный гликолиз. Промежуточные продукты окислительного распада глюкозы используются при синтезе аминокислот, липидов и других биомолекул. В наибольшей зависимости от аэробного распада глюкозы находится мозг. Он расходует около 100 г глюкозы в сутки. При голодании для организма очень важен процесс глюконеогенеза. Существует и прямой аэробный путь превращения глюкозы ‑ пентозофосфатный цикл, выполняющий анаболическую функцию поставки НАДФН для восстановительных синтезов и пентоз для синтеза нуклеотидов.
Цель:
1. Изучить гликолиз и глюконеогенез, гликогенолиз, спиртовое брожение, а также этапы аэробного распада углеводов и пентозофосфатного пути.
2. Изучить регуляцию распада и синтеза глюкозы, гликогена, нарушения обмена углеводов.
3. Ознакомиться с методами оценки гликолиза по обнаружению лактата, оценки содержания глюкозы в крови и моче и построения гликемических кривых.
Вопросы для самоподготовки
1. Важнейшие пути превращения глюкозы в тканях, роль глюкозо‑6‑фосфата.
2. Гликолиз: локализация, реакции (обратимые и необратимые, сопряженные с потреблением и синтезом АТФ), ферменты и метаболиты, суммарное уравнение, энергетический эффект. Суть субстратного фосфорилирования, роль.
3. Судьба восстановленного НАД, образовавшегося при окислении глицеральдегид‑3‑фосфата. Гликолитическая оксиредукция, ее сущность и значение.
4. Гликогенолиз, ферменты, суммарное уравнение, энергоэффект окисления глюкозы при распаде гликогена и глюкозы, поступающей из кровотока, сходство и отличие от гликолиза. Гликогенолиз с целью повышения концентрации глюкозы в крови, его энергетический эффект, локализация.
5. Метаболизм фруктозы и галактозы. Основы фруктозурии и галактоземии.
6. Спиртовое брожение, суммарное уравнение, энергетический эффект, сходство и отличие от гликолиза. Влияние этанола на обмен углеводов, причины гиперлактатемии и гипогликемии при острой интоксикации.
7. Предшественники глюкозы в глюконеогенезе, специфические реакции, последовательность общих реакций, ферменты, локализация процесса. Суммарное уравнение. Расход энергии для синтеза 1 молекулы глюкозы.
8. Регуляторные ферменты гликолиза и глюконеогенеза, аллостерические эффекторы, влияние гормонов. Глюкозолактатный и глюкозоаланиновый циклы, значение при длительной физической работе и голодании.
9. Направления окислительного распада глюкозы, специфические и общие пути. Реакции аэробного распада: а) окисление до пирувата, б) окислительное декарбоксилирование пирувата, в) окисление ацетил~КоА в ЦТК. Указать ферменты, коферменты, акцепторы Н, локализацию, суммарное уравнение.
10. Переносчики водорода глюкозы в дыхательную цепь. Окислительное фосфорилирование, сходство и отличия с субстратным фосфорилированием.
11. Выход АТФ при аэробном и анаэробном распаде глюкозы, сходство и отличия. Роль глицерол–фосфатного, малат–аспартатного челночных механизмов. Роль аэробного распада глюкозы в мозге, эритроцитах, при работе мышц. Биохимический механизм эффекта Пастера.
12. Пентозофосфатный путь окисления глюкозы: локализация и роль, реакции окислительного этапа и смысл неокислительного, ферменты и коферменты, связь с гликолизом, понятие о метаболической восстановительной энергии.
13. Нервная и гормональная регуляция обмена углеводов. Инсулин и контринсулярные гормоны, их влияние на процессы превращения углеводов.
14. Концентрация глюкозы в крови и ее источники. Гипо- и гипергликемии, их возможные причины. Нарушения обмена углеводов при сахарном диабете. Гликемические кривые, диагностическое значение, коэффициент Бодуэна. Изменения углеводного обмена при гипоксических состояниях.
Самостоятельная работа
Дополнить схему углеводно-энергетического обмена (темы 3.1 и 4.1). По теме 4.2 на схеме представить химизм и взаимосвязи основных метаболических путей обмена глюкозы – гликолиз и глюконеогенез, пентозофосфатный цикл, их выход на общие пути катаболизма (ПДГ-комплекс и ЦТК с энергетической цепью) для создания целостного представления об углеводно-энергетических потоках в организме. Приложить суммарные реакции показанных путей обмена.
Лабораторная работа 1.
Обнаружение молочной кислоты в биологических пробах
Лактат является конечным продуктом гликолиза и гликогенолиза, протекающих в анаэробных условиях, которые возникают при мышечной нагрузке как физиологического, так и патологического характера (приступ эпилепсии, столбняк, тетания и другие судорожные состояния); гипоксии, связанной с сердечной и легочной недостаточностью; анемии и других нарушениях. При снижении кислотности желудочного сока (гипоацидный гастрит и др. заболевания) снижаются его бактерицидные свойства, поэтому анаэробная микрофлора, попадающая из полости рта в желудок, оказывается в оптимальных условиях и нарабатывает лактат в процессе молочнокислого брожения углеводов пищи.
Принцип
Метод основан на реакции Уффельмана: при взаимодействии с лактатом комплексное соединение фенолята железа (III) фиолетового цвета превращается в малодиссоциирующую соль лактата железа, имеющую зеленовато‑желтую окраску.
Реактивы
1) Фосфатно-соляной буфер, рН 7,2, 2) 1 % раствор фенола, 3) 1 % раствор FeCl3, 4) 4 % раствор молочной кислоты.
обнаружение лактата в мышечной ткани
Материал для исследования
Мышечная кашица.
Проведение анализа
Готовят экстракт: кусочек мышцы тщательно растирают в ступке с 5,0 мл фосфатного буфера, полученную кашицу фильтруют через 2 слоя марли. Проводят цветную реакцию на лактат: в пробирке готорят раствор фенолята железа, добавляя к 10 каплям раствора фенола 1‑2 капли раствора FeCl3 до появления фиолетовой окраски, к содержимому пробирки добавляют 3 капли экстракта мышц и наблюдают изменение окраски. В присутствии лактата фиолетовая окраска переходит в зеленовато‑желтую вследствие образования лактата железа.
обнаружение лактата в желудочном соке
Материал для исследования
Патологический желудочный сок.
Проведение анализа
В пробирке готовят р-р фенолята железа из 2,0 мл р-ра фенола и 3 капель р-ра FeCl3, смесь разливают поровну в 2 пробирки. Затем в 1-ю пробирку добавляют 1-2 капли р-ра лактата (контроль), во 2-ю ‑ патологический желудочный сок. В присутствии лактата фиолетовая окраска заменяется зеленовато-желтой.
Оформление работы
Указывают принцип метода, регистрируют результаты в таблице, делают вывод: о путях снабжения мышцы кислородом, о присутствии молочной кислоты и активности анаэробной микрофлоры в образце желудочного сока.
| Исследуемая проба | Результат |
| Экстракт мышечной ткани | |
| Патологический желудочный сок |
Практическое значение
Наличие патологической кислотности (продуктов брожения: молочной, уксусной, масляной кислот) в желудочном соке указывает на сдвиг показателя кислотности от нормы в сторону повышения рН и появление в желудке анаэробных микроорганизмов и продуктов их жизнедеятельности.
Лабораторная работа 2.
Определение содержания глюкозы в сыворотке крови
Благодаря наличию сложных механизмов регуляции, включающих центральную нервную, эндокринную системы и деятельность печени, содержание глюкозы в крови здорового человека довольно постоянно и колеблется от 3,3 до 5,8 ммоль/л. Значительные отклонения в сторону увеличения содержания глюкозы называют гипергликемией (гиперглюкоземией), в сторону уменьшения ‑ гипогликемией (гипоглюкоземией). Для оценки содержания глюкозы в крови используют различные методы (редуктометрические, колориметрические, электрохимические, ферментативные), среди которых унифицированным является глюкозооксидазный. Метод специфичен, его можно использовать не только для оценки содержания глюкозы в плазме, сыворотке и цельной крови, но и в спинномозговой жидкости, транссудатах, экссудатах.
Принцип
Глюкоза с помощью глюкозооксидазы (КФ 1.1.3.4.) окисляется до глюконовой кислоты с образованием пероксида водорода. Пероксид водорода в присутствии фермента пероксидазы (КФ 1.11.1.17.) окисляет краситель 4-амино-антипирин, превращая его в окрашенное соединение. Интенсивность окраски пропорциональна содержанию глюкозы и определяется фотоколориметрически.
Материал для исследования
Сыворотка крови.
Реактивы
1) Рабочий реагент (фенол, 4-амино-антипирин, глюкозооксидаза, пероксидаза, в калиево‑фосфатном буфере), 2) 5,55 ммоль/л стандартный раствор глюкозы.
Проведение анализа
Готовят опытную и стандартную пробы согласно таблице
| Опыт, мл | Стандарт, мл | |
| Сыворотка Стандарт глюкозы Рабочий реагент | 0,01 ‑ 3,0 | ‑ 0,03 3,0 |
| Инкубируют 15 мин при 37°С. Измеряют оптическую плотность проб при длине волны 510‑530 нм в кювете 1 см против рабочего реагента. | ||
Расчет
Глюкоза [ммоль/л] =  ,
,
где: ЕОП – оптическая плотность пробы, ЕСТ – оптическая плотность стандарта, ССТ – концентрация стандартного раствора.
Нормальные величины
Цельная кровь 3,9‑5,8 ммоль/л
Сыворотка 3,3‑5,8 ммоль/л
Ликвор взрослые 2,75‑3,85 ммоль/л
Оформление работы
Указывают принцип метода, регистрируют результаты анализа, выполняют расчеты. Сравнивают полученные данные с нормальными значениями. Делают заключение о наличии патологических отклонений.
Практическое значение
Увеличение содержания глюкозы в крови свыше 6,0 ммоль/л наблюдается как при физиологических, так и при патологических состояниях.
К физиологической гипергликемии относят алиментарную (одномоментный прием больших количеств легкоусвояемых углеводов) и нейрогенную (выброс в кровь больших количеств катехоламинов при стрессе). Физиологические гипергликемии носят транзиторный характер и достаточно быстро проходят.
Патологическая гипергликемия, как правило, обусловлена нейроэндокринными расстройствами с нарушением соотношения между секрецией гормонов гипо- и гипергликемического действия. Наиболее частая причина ‑ сахарный диабет с абсолютной или относительной инсулиновой недостаточностью. Гипергликемия развивается при заболеваниях гипофиза с ростом секреции соматотропина и кортикотропина (акромегалия, болезнь Иценко–Кушинга, опухоли гипофиза и др.), опухолях мозгового слоя надпочечников с усиленным синтезом катехоламинов (феохромоцитома) и коркового слоя надпочечников с усиленной продукцией глюкокортикоидов, гиперфункции щитовидной железы, поражениях диэнцефальной области, болезнях печени (инфекционный гепатит, цирроз).
Физиологическая гипогликемия: алиментарная при недостатке поступления углеводов с пищей и голодании, при усиленном образовании и выбросе в кровь инсулина (ответ на алиментарную гипергликемию) – развивается при некомпенсированном потреблении углеводов как источника энергии, или после тяжелой и длительной мышечной работы. Может возникнуть в период лактации при усиленном поглощении глюкозы молочной железой.
Патологическая гипогликемия наблюдается при заболеваниях поджелудочной железы с гиперплазией b‑клеток островков Лангерганса ‑ гиперинсулинизм (инсулома, аденома, рак поджелудочной железы). Самая частая причина гипогликемий ‑ передозировка инсулина. Причиной гипогликемии может стать снижение продукции гормонов ‑ антагонистов инсулина при гипофункции коры надпочечников (Аддисонова болезнь, опухоли надпочечников), гипофункции и атрофии передней доли гипофиза (болезнь Симмондса), гипофункции щитовидной железы. Нейрогенная гипогликемия наблюдается при заболеваниях нервной системы (энцефалит, прогрессивный паралич и др.), психических заболеваниях (хронический алкоголизм, циклотемия и др.), травмах головного мозга. Гипогликемия может возникать при тяжелых поражениях печени (отравления фосфором, хлороформом, острая желтая дистрофия печени, цирроз и др.), гликогенозах (болезнь Гирке) вследствие невозможности превращения гликогена в глюкозу. Гипогликемия при заболеваниях почек обусловлена потерей значительного количества глюкозы с мочой вследствие снижения почечного порога для глюкозы. Гипогликемия наблюдается при врожденных дефектах жирового и углеводного обмена в связи с неспособностью организма эффективно мобилизовать свои энергетические ресурсы.
Лабораторная работа 3.
Влияние сахарной нагрузки на содержание глюкозы в крови
(глюкозотолерантный тест)
Метод сахарной нагрузки (глюкозотолерантный тест (ГТТ) или тест на толерантность к глюкозе (ТТГ)) информативен для выявления скрытой формы сахарного диабета и нарушения гликогенообразовательной функции печени.
Принцип
Метод основан на определении содержания глюкозы в крови до нагрузки глюкозой и через 30, 60 и 120 мин после проведения нагрузки. Содержание глюкозы в крови определяют глюкозооксидазным методом (см. предыдущую работу).
Материал для исследования
Образцы капиллярной крови, взятой до нагрузки глюкозой и через определенные промежутки времени после нагрузки.
Проведение анализа
В клинических лабораториях при проведении пробы с сахарной нагрузкой у обследуемого натощак берут кровь из пальца, определяют в ней содержание глюкозы. Затем дают раствор глюкозы или сахарозы из расчета 1,0‑1,5 г сахара на 1 кг массы тела. Через 30, 60 и 120 мин после приема сахара снова берут образцы крови и в них определяют содержание глюкозы.
На практическом занятии сахарную нагрузку изучают с готовыми образцами крови, взятыми до нагрузки глюкозой, через 30, 60 и 120 мин после нее.
Готовят опытные и стандартную пробы согласно таблице
|
| Пробы, мл | Стандарт, мл | |||
| до | время после нагрузки | ||||
| 30 мин | 60 мин | 120 мин | |||
| 1 | 2 | 3 | 4 | 5 | |
| Рабочий раствор Кровь Стандарт глюкозы | 3,0 0,01 ‑ | 3,0 0,01 ‑ | 3,0 0,01 ‑ | 3,0 0,01 ‑ | 3,0 ‑ 0,01 |
| Содержимое проб перемешивают, инкубируют 15 мин при 37°С, измеряют экстинкцию при длине волны 510‑530 нм в кювете 0,5 см против рабочего раствора, рассчитывают концентрацию глюкозы. | |||||
Расчет
Глюкоза [ммоль/л] =  ,
,
где: ЕОП ‑ оптическая плотность соответствующих опытных проб (до нагрузки, через 30, 60 и 120 минут после неё), ЕСТ ‑ оптическая плоность стандарта, ССТ ‑ концентрация стандартного раствора.
На основании полученных данных строят график ‑ гликемическую (сахарную) кривую, откладывая по оси абцисс время взятия крови, по оси ординат ‑ содержание глюкозы в соответствующей пробе.
Нормальные величины
натощак 3,9‑5,8 ммоль/л (100 %)
через 60 мин 6,7‑9,4 ммоль/л (150‑175%)
через 120 мин ниже 6,7 ммоль/л
Практическое значение
Уровень глюкозы в крови у здоровых лиц после нагрузки глюкозой:
1. Через 30‑60 мин после приема глюкозы наблюдается максимальное увеличение содержания глюкозы в крови: на 35‑80% выше исходного. Повышение содержания глюкозы в течение 1-го часа после ее приема объясняется переходом ее в кровь и в значительной мере определяется быстротой всасывания, гликогенсинтезирующей функцией печени и всех прочих периферических органов.
2. Через 90‑120 мин содержание глюкозы в крови возвращается к норме и в ряде случаев может быть даже ниже исходной величины. Уровень глюкозы в крови в этот период снижается из-за усиленного выделения инсулина поджелудочной железой в ответ на развивающуюся гипергликемию, а глюкоза переходит в ткани. Инсулина выделяется больше, чем требуется для восстановления нормального уровня глюкозы в крови, что приводит к небольшой гипогликемии.
3. К 150‑180 мин содержание глюкозы в крови возвращается к исходному за счёт достижения равновесия всех систем регуляции уровня глюкозы в крови.
В клинической практике выделяют несколько видов гликемических кривых – нормальная, гипергликемическая и гипогликемическая:
| Параметры по содержанию глюкозы | Кривые | ||
| Нормальная | Гипергликемическая | Гипогликемическая | |
| 1. Исходный уровень | норма | гипергликемия | гипогликемия |
| 2. Максимальный подъем | 1 ч | 2‑3 ч | Замедлен на 2‑3 ч |
| 3. Фаза гипогликемии | 2 ч | нет | Резко выражена |
| 4. Уровень к концу третьего часа | Исходный уровень | Исходного уровня не достигает | Исходного уровня не достигает |
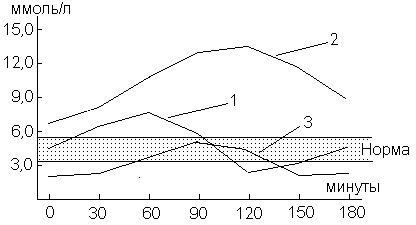
Типы гликемических кривых:
Дата добавления: 2020-04-25; просмотров: 569; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!