Ветеринарно-профілактичні загальні заходи при виникненні абортів
Кожен випадок аборту необхідно детально вивчити, з’ясувати причину і прийняти необхідні заходи. До виключення інфекційної причини аборту він розцінюється як заразне захворювання і в господарстві проводять заходи, передбачені ветеринарним законодавством.
Оскільки більшість абортів викликаються цілим комплексом порушень взаємо-відношення між материнським організмом та плодом, тому в кожному випадку слід встановити головну ланку цих порушень.
Конкретну причину аборту іноді важко встановити, оскільки смерть плода не за-вжди наступає зразу після дії абортогенного фактора. В окремих випадках від смерті плода до його зігнання може пройти значний проміжок часу.
В кожному окремому випадку необхідно перш за все:
Ø виключити заразний характер аборту (на підставі комплексного врахування епізоотичної ситуації господарства; даних клінічного обстеження тварини, що абор-тувала; лабораторного дослідження матеріалу від тварин, що абортували, та закріпле-них за ними плідників);
Ø виключити ідіопатичний незаразний аборт на ґрунті аномалій розвитку пло-дових оболонок чи патологічних процесів у них (досліджуючи оболонки та плід);
Ø виключити аліментарні аборти (на підставі дослідження кормів та умов годів-лі тварин);
Ø виключити можливість травматичного аборту;
Ø виключити симптоматичні аборти на ґрунті (заразних чи незаразних) захво-рювань.
Висновок можна робити лише на підставі комплексного дослідження абортова-ного плода, його оболонок, материнського організму з обов’язковим аналізом умов догляду, утримання та експлуатації.
Оскільки після аборту у тварини можуть спостерігатися різноманітні ускладнення (атонія матки, ендометрит, затримання посліду та ін.) із загальною реакцією організ-му, тому вживають заходи щодо їх лікування та профілактики.
Патологія вагітності
Питання для самоконтролю
1. Які патологічні процеси спостерігаються у вагітних самок?
2. Яку лікувальну допомогу слід надати тварині при передчасних переймах та по-тугах?
3. Яку допомогу необхідно надати вагітній корові при її залежуванні?
4. Які розрізняють види випадіння піхви у корів і яку допомогу надають при цьому? 5. Що розуміють під додатковою та позаматковою вагітністю?
6. Згадайте класифікацію абортів за А. П. Студєнцовим. 7. Які причини ранньої ембріональної смертності?
8. Які основні ознаки муміфікації плода і яку допомогу слід надати тварині? 9. Які основні ознаки мацерації плода і яку допомогу слід надати тварині? 10. Які ознаки путрифікації плода і яку допомогу слід надати тварині?
11. Які причини ідіопатичних незаразних абортів?
12. Які причини ідіопатичних інфекційних та інвазійних абортів? 13. Які основні причини симптоматичних незаразних абортів?
14. Які основні причини симптоматичних інфекційних та інвазійних абортів? 15. Які заходи проводять у господарстві при виникненні абортів у тварин?
| 10. |
10.1. Теорії початку родового процесу
Роди (Рartus) – складний фізіологічний процес, суть якого полягає у виведенні з організму матері дозрілого(их) і живого(их) плода(ів), звільненні порожнини мат-ки від плодових оболонок та залишків плідних рідин у характерні для кожного виду тварин терміни. Не дивлячись на велику кількість проведених наукових досліджень цей фізіологічний процес остаточно не вивчений. Загадковими і нерозшифрованими як для вчених, так і для практиків залишається питання: “Чому роди у більшості ви-падків відбуваються вночі або зранку, а не в день?”; “Чому впродовж усього плодоно-шення самка зберігає плід і лише вкінці його розпочинається родовий процес?”; “Що є стартовим механізмом початку родового процесу – реакція нервової, гуморальної чи іншої систем, які є відповідальними за цей складний механізм?”, “Чому різноманітні фактори зовнішнього або ж внутрішнього середовища в одних екстремальних умовах здатні прискорити, в інших – загальмовувати, а ще в інших не спричинити ніякого впливу на цей складний фізіологічний процес?”, “Чому фактор годівлі самок за 1–2 доби до умовного терміну родів володіє синхронізуючою дією для одночасного про-яву цього процесу у багатьох вагітних самок?” та багато інших питань, над якими ще не один десяток років будуть працювати вчені-акушери.
Більшість учених, які займалися вивченням фізіології родів, схиляються до дум-ки, що під час них відбувається цілеспрямована переорієнтація нейроендокринної ре-гуляції статевої системи самок на виведення плода з утроби матері.
Причину родів вбачають також у дегенеративних процесах в оболонках плода, і головним чином у плаценті, які порушують зв’язок між плодом і матір’ю, що веде до зігнання плода з порожнини матки, як стороннього тіла.
Початок родової діяльності пояснюють і впливом гормональних факторів. З розвитком вагітності в організмі самки збільшується кількість секретів надниркових залоз та задньої частки гіпофіза, які мають здатність спричиняти скорочення матки. Уже на 9-му місяці вагітності корів жовте тіло припиняє виділення прогестерону і в яєчнику починає рости фолікул, який синтезує естрогени. Під їх впливомвідбувається
Фізіологія родів
розрідження і витікання слизового корка шийки матки таїї відкриття. Зміна маткового тиску спричиняє скорочення міометрію і початок родів.
Впродовж тривалої історії вивчення механізмів родів були висловлені різні теорії і гіпотези, але жодна з них в повній мірі не може пояснити і розкрити причини цього складного фізіологічного процесу, хоча, ніде правди діти, жодна з них не може бути відкинута і в якійсь мірі заслуговує на увагу. В кожній з цих теорій автори наводять причину, яка на їх думку є стартовою, першочерговою і яка викликає перше скоро-чення м’язів матки. Серед них можна назвати теорії: тиску, розтягнення м’язів матки, стороннього тіла, токсикозу, подразнення нервових закінчень м’язів матки вуглекис-лотою, теорію зрілості м’язів матки, реакції маткового центру на подразнення та не-йрогуморальну теорію Ніколаєва – Лазарєва – Петченка.
Теорія тиску. Окремі автори вважають, що плід, досягнувши свого зрілого стану, сильно тисне на тканини матки, в яких міститься велика кількість нервових гангліїв, особливо в ділянці дорсальної частини шийки матки, викликаючи при цьому рефлек-торне скорочення м’язів матки.
Теорія розтягнення. Автори цієї теорії вважають, що в кінці вагітності плід, що інтенсивно росте, розвивається, заповнює порожнину матки і призводить до макси-мального розтягнення її стінок, що є подразником для початку скорочення м’язів і родового процесу.
Теорія стороннього тіла. Причиною початку родів, на думку авторів, є дегене-ративні зміни та переродження і процеси в оболонках плода (плодів), які порушують зв’язок між плодовою і материнською частинами плаценти і плід виявляється сторон-нім тілом, що обумовлює скорочення матки та виведення його з її порожнини.
Теорія токсикозу. Автори даної теорії пояснюють початок родової активності по-стійним накопиченням в організмі матері великої кількості продуктів обміну речовин, що надходять як від самого плода, так і з плаценти, і які є фактичними подразниками хеморецепторів матки, викликаючи її збудження і початок скорочення.
На думку авторів теорії реакції маткового центру в живому організмі людини і тварини всі без винятку життєво важливі фізіологічні процеси, в тому числі й розмно-ження, контролюються і регулюються відповідними центрами центральної нервової системи, тобто родовий процес регулюється через так званий “матковий центр”. Із збільшенням строків вагітності, особливо в її кінці, в організмі матері накопичують-ся гормони наднирників, задньої долі гіпофізу, плаценти, що обумовлює подразнен-ня механо-хемо-барорецепторів матки, а через них і відповідних маткових центрів у головному і спинному мозку. Тут ці імпульси концентруються, нагромаджуються і викликають формування родової домінанти. Після досягнення певного рівня вони викликають перші родові перейми у вигляді потужного скорочення матки, що є по-чатком родового акту.
Розділ 10
Нейрогуморальна теорія Ніколаєва – Лазарєва – Петченка. Автори даної тео-рії розділили всі причини початку родів на три групи: а) фактори, що підготовляють роди; б) фактори, що викликають роди; в) фактори, що підтримують ритміку родів.
Фактори, що підготовляють роди: а) зниження збудливості кори і підвищення збудливості спинногомозку; б) підвищення збудливості нервово-м’язового апаратумат-ки; в) підсилення подразнень баро- і механорецепторів матки; г) тонотропна дія адре-нергічної іннервації матки; д) накопичення естрогенів і зменшення концентрації прогес-терону; є) накопичення глікогену і глютатіону, які підвищують збудливість міометрію.
Фактори, що викликають роди: а) тономоторна дія холінергічних механізмів матки; б) подразнення рецепторів матки і її шийки плодом, формування родової домі-нанти; в) розвиток гормонального “поштовху” на матку (різке зростання концентрації естрогенів).
Фактори, що підтримують роди: а) швидкий тономоторний ефект ацетилхоліну; б) нейтралізація (руйнування) холінестеразою ацетилхоліну; в) гальмівна дія оксито-цину (пітоцину) на холінестеразу; в) антагоністична (пригнічуюча) дія пітоцинази на окситоцин; г) гальмівна дія естрогенів на пітоциназу; д) подразнення парацервікаль-них та паравагінальних гангліїв, які викликають скорочення матки.
Згідно з цією теорією, родова діяльність розпочинається внаслідок подразнення ацетилхоліном, концентрація якого в кінці вагітності у плаценті різко зростає, ганглі-їв, розміщених у нервових сплетіннях матки. Ацетилхолін постійно накопичується в плаценті і в кінці вагітності його концентрація максимальна. Крім цього, в кінці вагітності в крові матері накопичуються у великій кількості такі гормони, як окси-тоцин, адреналін та естрогени, які також стимулюють початок родової діяльності. Одночасно з цим, початок родової діяльності забезпечується значним зниженням в кінці вагітності вмісту в крові такого гормону, як прогестерон, безпосередня дія яко-го направлена на збереження спокою вагітної матки, тобто на гальмування її скорот-ливої здатності.
Тобто, родовий акт є результатом складного комплексу взаємодії фізіологічних процесів, які відбуваються в материнському організмі протягом тривалого часу.
Через привідкритий цервікальний канал у порожнину матки входить повітря, змі-нюється внутрішньоматковий тиск, починаються перейми, що викликає рух плода і тиск на стінку та шийку матки, тобто це і можна вважати початком родів (С. П. Хо-мин, 1985).
Тобто, головним подразником, який викликає початок родів, є плід, який своїм тиском і рухами подразнює нервові закінчення (рецептори) матки. Безпосередньо пе-ред родами під прямим впливом релаксину та непрямим впливом інших гуморальних факторів відбувається розслаблення зв’язок кісткового тазу і розкриття шийки матки.
Отже, початок і перебіг родів обумовлені впливом таких факторів:
Нервово-рефлекторні фактори – зниження в кінці вагітності збудливості кори головного мозку і одночасне зростання збудливості спинного мозку та рецепторів матки; утворення в корі головного мозку “родової домінанти”; підвищення чутли-
Фізіологія родів
вості (сенсибілізації) нервово-м’язових структур матки до окситоцину і інших біо-логічно активних речовин локомоторної дії; часткова “фізіологічна десенсибілізація” чутливих інтрамуральних нервових утворень (рецепторів) матки щодо залишків, що утворилися під час вагітності.
Гормональні фактори – зростання синтезу в кінці вагітності і на початку родів фето-плацентарним комплексом і яєчниками естрогенів (естрадіолу, естрону і, осо-бливо, естріолу), що мають багатогранний вплив на нервово-м’язові елементи матки. При цьому зростає збудливість, провідність та реактивність міометрію до біологічно активних речовин: окситоцину, простагландинів, серотоніну, ацетилхоліну, катехола-мінів і кінінів, а також до інших аналогів окситичної дії. Зміна співвідношення поміж естрогенами і прогестероном у бік переваги естрогенів веде до зняття прогестероно-вого блоку міометрію, а також до втрати прогестероном, а можливо й хоріонічним гонадотропіном (ХГ) і хоріонічним соматотропіном антиконтрактильних властивос-тей; підвищення зростання у крові в кінці вагітності рівня кортикостероїдів, які тісно зв’язані з естрогенами та необхідні для забезпечення скорочувальної функції матки.
Нейрогуморальні фактори – підсилення в кінці вагітності і перед родами син-тезу окситоцину, серотоніну, простагландинів, катехоламінів, ацетилхоліну, кінінів та різкий спад активності ферментів, що їх нейтралізують – окситоцинази (пітоцинази), холінестерази, моноамінооксидази, кінінази та ферменту, який нейтралізує проста-гландини; зміна чутливості міометрію до біологічно-активних речовин (підвищення чутливості α-рецепторів і зниження активності β-рецепторів).
Біоенергетичні фактори, які визначають готовність матки в кінці вагітності до посилення скорочення міометрію – зростання синтезу в кінці вагітності актоміозину (скорочувального білка), глікогену, АТФ та інших фосфорних з’єднань, електролітів (кальцію, натрію, калію), мікроелементів (кобальту, заліза, цинку).
Метаболічні і трофічні фактори – велике накопичення деяких продуктів обміну речовин плода (компенсований ацидоз плодів веде до посилення їх рухів); високий прояв дегенеративних процесів, що відбуваються у дозрілій плаценті, повний роз-виток м’язових елементів, еластичних й інших сітчасто-волокнистих сполучнотка-нинних структур матки; функціональна зрілість контрактильної системи міометрію (міофібрил, актоміозину); максимальна функціональна активність білків саркоплазми м’язових клітин матки та їх субклітинних субстанцій – мітохондрій та рибосом, і на цьому фоні – різке підвищення інтенсивності та енергетичної ефективності окислю-вальних процесів (зростання активності сукцинатдегідрогенази, цитохромоксидази, збільшення тиску кисню та інтенсивності його засвоєння тканинами матки).
Механічні фактори – призупинення вільного розтягнення матки в результаті зни-ження активності прогестерону, значне збільшення і остаточне дозрівання плодів та посилення їх рухової активності; розсмоктування та різке зменшення продукції ам-ніотичної рідини і максимальне приближення першого плоду до шийки матки, що також викликає механічне подразнення нервових гангліїв сенсибілізованих естроге-нами рецепторів матки, шийки матки звільнення її від слизового корка та відкриття.
Розділ 10
10.2. Зміни матки під час родів
Фізіологічна активність матки визначається перш за все функціональним станом її нервово-м’язових структур, основними параметрами чого є тонус, збудливість, провідність, скоротливість та реактивність на біологічно активні речовини. З набли-женням строків родів тут домінують α-рецептори. Зростання концентрації естроге-нів з паралельним зниженням вмісту прогестерону і викликає ослаблення чутливості β-рецепторів, що обумовлює скорочення матки. Тому блокада β-рецепторів має прак-тичне значення при збудженні родової діяльності.
Стимуляція α-адренергічних рецепторів матки підсилює її скорочення, в той час, як їх блокування – знижує їх. Стимуляція β-рецепторів знижує скорочувальну актив-ність матки.
В процесі родів розтягнуті м’язові волокна матки при кожній переймі все більше скорочуються (контракція), взаємно зміщуються, переплітаючись одне з одним в ре-зультаті чого відбувається їх так звана ретракція (нашарування). З ретракційним пере-групуванням м’язових волокон матки тісно пов’язана дистракція (розтягнення) ший-ки матки. Іншими словами, в процесі родів відіграють роль три фактори – контракція, ретракція і дистракція м’язових волокон, а також процес активного їх розслаблення.
При цьому спрацьовує феномен “домінанти верхівки рогу”, суть якої полягає в тому, що м’язи верхівки рогів матки починають скорочуватися раніше, більш інтенсив-но і протяжно, ніж інші сегменти матки. При нормальних родах хвиля скорочення роз-повсюджується впродовж 30–80 сек. від кінцевих ділянок яйцепроводів до тіла матки.
Що стосується місця виникнення першого – пускового скорочення матки, за яким іде розповсюдження скорочувальної хвилі, тотут існує декілька думок. За аналогією з діяльністю м’язів ряд авторів висловлюються за наявність у міометрії ділянок так зва-ного “водія ритму”, де зароджуються скорочення. З іншого боку, інші вчені вважають, що в матці немає якогось центру зародження скорочення і шляху розповсюдження імпульсу. Кожний сегмент міометрію може функціонувати як синусний вузол. Кожна точка м’язів матки може стати спонтанно активною і відіграти роль синусного вузла або “водія ритму”. Розміщення цієї місцевої активності залежить лише від йонної рівноваги у м’язовій тканині.
З точки зору йонної теорії збудження, в основі скорочувальної активності матки при родах лежать зміни мембранних потенціалів м’язової клітини.
Властивість живої тканини збуджуватися, тобто відповідати на зовнішні подраз-ники тісно пов’язана з тонусом маткових м’язів. Тонус і збуджуваність матки визна-чаються морфо-функціональним станом нервово-м’язових структур.
Значній морфологічній і функціональній перебудові під час вагітності і родів під-дається і судинна система матки. Кровоносні судини, особливо вени, стають довши-ми, стінки судин гіпертрофуються, кровонаповнення значно підсилюється. Швидкість кровотоку у матці в кінці вагітності прискорюється, насиченість крові киснем зростає в декілька разів.
Фізіологія родів
Перейми і потуги ведуть до зменшення об’єму циркулюючої крові в міометрії, а це викликає тимчасову його гіпоксію, що особливо виражене в кінці періоду розкрит-тя шийки матки (підготовчого).
При нормальних родах в період пауз між переймами і потугами гемодинаміка в міометрії швидко відновлюється, а при слабкості родової діяльності відбуваються значні відхилення в гемодинаміці – утруднений приплив і відтік крові як результат зменшення судинного тонусу матки, інколи спазматичні явища.
Відомо, що однією з умов розвитку родового процесу є оптимальний стан вну-трішнього середовища організму роділлі, причому, особливе значення має стан кис-лотно-лужної рівноваги крові. У вагітних перед родами в крові відзначається компен-саторний метаболічний ацидоз та дихальний алкалоз. Під час першого і, особливо, другого періоду родів метаболічний ацидоз підсилюється, але, як правило, залиша-ється компенсованим.
Родові шляхи
Таз як шлях для народження плоду. Родові шляхи складаються з двох основних частин: 1) кістково-зв’язової основи і 2) м’яких частин.
Кістково-зв’язовою основою родових шляхів є таз, який складається з двох без-іменних кісток, що з’єднуються одна з одною своїми нижніми частинами за допомо-гою тазового зрощення. Кожна з безіменних кісток в свою чергу складається з трьох кісток: широкої клубової кістки, що утворює суглоб з крилами крижів, лобкової і сід-ничної кісток, що утворюють дно таза. У новонароджених тварин усі три кістки спо-лучені хрящем, а у дорослих вони зростаються у одну кістку.
Крім клубових кісток, тазова порожнина з боків обмежена ще широкими тазови-ми зв’язками; зверху – крижовою кісткою і першими хвостовими хребцями; спере-ду – вона безпосередньо межує з черевною порожниною і нарешті, ззаду обмежена шкірно-м’язовою стінкою, так званою діафрагмою таза.
Кістковий таз є міцною основою для м’яких родових шляхів.
Широкі тазові зв’язки, що замикають порожнину таза з боків, мають у процесі родів важливе значення. Перед початком родів вони набрякають, розслаблюються і стають податливими під час проходження плода з матки. Зв’язки, що з’єднують крижі з тазом, теж під кінець вагітності розслаблюються, що створює можливість деякого переміщення крижів щодо таза. В результаті цього збільшується висота входу в таз і трохи розширюється простір між клубовими кістками.
Діафрагма таза має своєю основою м’язи і фасції. У верхній частині через діафрагму проходить пряма кишка, а у нижній – піхва. Пружність діафрагми зу-мовлює звуження піхви і є іноді перешкодою для проходження широких ділянок плода. Надмірне розслаблення м’язів діафрагми є фактором, що сприяє випадін-ню піхви назовні.
 Розділ 10
Розділ 10
1 13 14 13
2 123
1 18
Для встановлення роз-мірів (об’єму) тазу користу-ються пельвіометрією – ви-значенням промірів тазу, що дозволяє констатувати
7 А
8 1511 16
17 Б
параметри входу в таз, ви-ходу з тазової порожнини, характер тазової осі, без чого не можливо обрати ра-ціональну техніку родової допомоги і уникнути мож-ливих ускладнень.
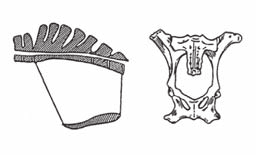
Рис. 77. Проміри тазу кобили (А) та корови (Б, вигляд спереду):
1 – крижова кістка; 2 – кінець тіла крижової кістки; 3 – хвостовий хребець; 4 – виступ крижової кістки; 5 – висота входу в таз; 6 – вертикальний діа-метр входу в таз; 7 – краніальний край симфізу лобкових кісток; 8 – ко-сий діаметр тазової порожнини; 9 – висота виходу з тазу; 10 – каудальний край симфізу сідничних кісток; 11 – вертикальний діаметр виходу з тазу; 12 – маклаки; 13 – крижово-клубні з’єднання; 14 – верхній діаметр входу в таз; 15 – поперечний діаметр тазової порожнини; 16 – западинна части-на лобкової кістки; 17 – нижній діаметр виходу з тазу; 18 – верхній діа-метр входу в таз; 19 – поперекові горбики; 20 – середній діаметр входу в таз; 21 – сіднична кістка; 22 – нижній діаметр входу в тазову порожнину;
23 – сідничні горби.
У практиці ветеринар-ного акушерства користу-ються такими параметрами тазу: верхній, середній та нижній поперечний діаме-три входу в таз; поперечний діаметр тазової порожни-ни; поперечні діаметри ви-ходу з таза; вертикальний діаметр таза, вертикальний діаметр тазової порожнини, довжина таза, тазова вісь.
Для практики акушера важливі тільки названі нижче проміри:
– тазова вісь – поздовжня лінія, що проходить через центр тазової порожнини на однаковій відстані від усіх однойменних кісткових виступів, тобто вектор прохо-дження плода під час родів через тазову порожнину;
– висота входу в таз – лінія, проведена від переднього краю лобкового зрощен-ня до переднього кінця тіла крижів;
– вертикальний діаметр тазової порожнини – перпендикуляр, проведений від краніального краю лобкового зрощення до крижової кістки;
– середній поперечний діаметр тазової порожнини – віддаль між серединами сідничних гребенів (spina ischiadica) обох безіменних кісток.
Таз кобили має найсприятливішу будову для родів серед усіх інших видів свій-ських тварин, він доволі обширний. На бокових стінках таза розміщені широкі зв’яз-ки з товстим шаром м’язів, що забезпечує еластичність тазової порожнини. Вхід до таза майже округлий, тільки донизу має незначне звуження. Найбільша ширина – по верхньому поперечному діаметру. Дно таза майже горизонтальне, тільки передній край його трохи опущений, що збільшує розміри входу в таз. Вісь таза кобили має вигляд лінії, зверненої незначним викривленням догори.
 Фізіологія родів
Фізіологія родів
Крижі під час родів бувають мало-рухомі, бо зчленування їх з клубовою кісткою щільніше; це в процесі родів особливого значення немає, бо розміри тазової порожнини великі, а бокові стін-ки дуже податливі.
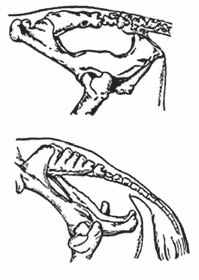
Таз корови має будову, несприятли-ву для проходження плода під час родів. Він має овальний вхід, витягнутий у до-вжину і форму зрізаного конуса.
Вісь таза має вигляд ламаної лінії. Дно таза жолобкувате і має великий підйом у задній частині, що спричиняє зміну напрямку руху плода під час ви-ходу з родових шляхів, а тому частини плода, що увійшли в тазову порожнину, можуть тут натрапляти на деякі пере-шкоди дальшому просуванні назовні.
Під тиском плода, що проходить по родових шляхах, крижі трохи зміщу-ються догори. Це зміщення можливе у корів, як наслідок, не зовсім міцного зчленування крижів з клубовою кіст-
А
Б
Рис. 78. Тазова вісь корови (А) і кобили (Б).
кою. Насильне натиснення крижової
кістки догори при витяганні занадто великого плода може спричиняти розтягнення зв’язок крижово-клубового зчленування, або навіть розрив їх, що призводить до важ-кого захворювання (залежування після родів).
Таз дрібних жуйних, свиней і м’ясоїдних має в анатомічному відношенні такі осо-бливості в будові тазової порожнини, що створюють умови, сприятливі для родів.
М’які структури родових шляхів представлені тканинами матки і шийки, піхви та вульви. Під час родів шийка матки повільно розкривається повністю і її межа між зовнішніми і внутрішніми отворами практично зникає. Піхва легко розтягується і не протидіє проходженню плоду. Розтяжність вульви дещо менша ніж піхви, тому великі плоди під час родів інколи затримуються в цьому місці.
Коли шийка матки цілком розкривається, межа між зовнішніми і внутрішніми отворами її згладжується. Тільки у корів іноді верхня стінка втулкоподібного виступу шийки матки спостерігається у вигляді складки в порожнині піхви.
У тазовій порожнині розташовані, крім піхви і шийки матки, ще пряма кишка і сечовий міхур. Через те, що пряма кишка і сечовий міхур на початку родів спо-рожняються, вони в цьому стані не перешкоджають розширенню м’яких родових частин.
Розділ 10
10.4. Передвісники родів та їх клінічні ознаки
У вагітних самок незадовго до настання родів проявляються характерні клінічні ознаки або симптомокомплекс, на підставі якого можна прогнозувати приблизно по-чаток родового процесу, але встановити точно цей час практично не можливо. У са-мок різних видів тварин вищезгаданий симптомокомплекс має свої особливості.
Кобила. В кінці вагітності спостерігається набряк і гіперемія статевих губ, за-падають крижі (чітко спостерігається западина поміж основою хвоста і сідничними горбами) і з’являється незначний набряк живота. Добре помітний набряк спостеріга-ється у кобил, які мали недостатню кількість прогулянок і активного моціону. Поряд з цими ознаками, відбувається помітне звисання живота під тиском плода на розпу-шені і м’які черевні стінки. Приблизно за 5–7 діб до родів молочна залоза кобили наповнюється молозивом, яке навколо отворів дійкових каналів засихає у вигляді ян-тарних крапель. Зменшення довжини шийки матки, яке виявляють при ректальному дослідженні кобил, свідчить про початок її розкриття. Короткочасним у кобил буває розрідження густого слизового корка шийки матки і його витікання за межі вульви у вигляді ниток тягучого слизу.
Корова. В кінці вагітності спостерігається звисання живота, особливо у тварин з неодноразовими родами. За 12–36 годин до початку виведення плода таз корови пе-ретворюється із “звичайного” в “родовий” за рахунок розслаблення його зв’язкової системи (зв’язки таза стають розпушеними і збільшуються на 1/3 – 1/4 їх довжини). Сідничо-крижова зв’язка, яка до родів при натискуванні на неї в ділянці підхвосто-вої складки пальпується у вигляді міцного непіддатливого тяжу, безпосередньо перед родами розслаблюється і її можна охопити пальцями та відтягнути на бік, легко змі-щуючи; в окремих випадках її контури згладжуються настільки, що вона взагалі не промацується. Западини між основою хвоста і сідничними горбами виразно збільшу-ються (“западання крижа”).
Іноді у окремих тварин зв’язки таза розслаблюються лише перед самим початком родів, або, навпаки, за 2–3 тижні до їх початку. Статеві губи збільшуються і набряка-ють, шкіра їх стає гладенькою і гіперемованою, складки розправляються.
За 1–2 доби до родів густий слизовий корок шийки матки та густий липучий слиз піхви розріджуються і виділяються за межі вульви у вигляді “поводків” чи тяжів в’яз-кого слизу, які звисають із зовнішніх статевих органів. Інколи аналогічна ситуація у 30–40 % корів може спостерігатися в кінці четвертого або на п’ятому місяці вагіт-ності, що більшістю авторів пояснюється різкими перебудовами і змінами в гормо-нальній системі самок, а точніше, включенням усистему гормоногенезусформованої плаценти. Малодосвідчені тваринники сприймають такі ознаки за прояви статевого циклу і навіть проводять осіменіння таких тварин, викликаючи аборт.
За 2–3 доби до родів, інколи й раніше, а в деяких випадках під час або навіть після родів вим’я наповнюється молозивом.
В останній місяць тільності корів простежується тенденція до незначного підви-щення загальної температури, але за 10–50 годин до початку родового процесу темпе-
Фізіологія родів
ратура тіла тварини може знижуватись на 0,4–1,2 ºС; за 2–3 години до початку родів зменшується довжина шийки матки, що можна виявити при ректальному дослідженні. За 1–2 доби до родів наступають відповідні зміни і в поведінці тварини: вини-
кає занепокоєння, вона оглядається на живіт, переступає з ноги на ногу, часто лягає і встає з підлоги, мукає, у неї знижується апетит, з’являється спрага.
Коза і вівця. За 3–4 тижні до родів починає збільшуватися вим’я, в останні 1–3 дні перед родами воно набрякає. Молозиво у молочній залозі з’являється за 1–2 доби, але найчастіше – безпосередньо перед родами, за 1–2 доби до окоту таз перетворю-ється на “родовий”.
За 3–4 доби до родів спостерігається гіперемія та припухання вульви, розріджен-ня слизового корка вагітності (пускання повідків); за 12–24 години до окоту кози чи вівці стають занепокоєними, часто гребуть підстилку, оглядаються на живіт, відмов-ляються від корму і жалібно мекають.
Свиноматка. У свині в кінці поросності відмічається висока ступінь звисання живота. За 10–12 годин до початку родів її таз перетворюється із звичайного в родо-вий за рахунок розслаблення крижово-сідничних зв’язок, втрачаючи випученість та округлість крупа у ділянці сідничних горбів. Зовнішні статеві органи піддаються гі-перемії і набрякають. За 1–2 доби до виведення плодів у свині розкривається шийка матки, а густий слизовий корок, який її закривав під час вагітності, розріджується і витікає за межі вульви у вигляді ниток тягучого слизу. За 24–48 годин до початку ро-дів у молочних залозах з’являється молозиво, у свиноматок, які мають світлу шкіру, відмічається почервоніння молочної залози.
Приблизно за добу, особливо за 6–12 годин до родів, проявляється так званий “ін-стинкт” наближення родів – свиноматка часто лягає і встає, риє підстилку, переносить її в роті, підготовлюючи у певному місці станка “гніздо”.
Сука. У цих тварин помічається теж обвисання живота, розслаблення тазових і крижових зв’язок, набряклість статевих губ і готування в затишному місці гнізда. Дуже характерною ознакою в кінці вагітності є зниження температури тіла. Воно по-чинається поступово, переважно за 2–3 дні до настання родів, але в окремих випадках зниження температури буває раптове. Проте завжди температура тіла за 24 години до настання родів досягає субнормальні межі (нижче 37 °С). Якщо врахувати, що в здо-рових собак нормальна температура коливається між 38–39 °С, то температура падає на 1–2 °С. З настанням потуг температура тіла швидко підвищується до норми.
Кішка. За тиждень до родів кішка починає часто вилизувати себе, особливо ді-лянку живота і зовнішніх статевих органів. Вона стає дуже роздратованою, а інколи й агресивною, особливо в присутності сторонніх осіб, шукає місця схованки для обла-штування свого гнізда, яким можуть бути шафи, темні місця, скриньки та інші меблі, де накопичений одяг або взуття, “перериває” господарське ліжко або диван, риє гніздо в саду чи допоміжних будівлях, в тому числі й сусідських, стає сильно збудженою.
За 10–30 годин до родів таз кішки перетворюється в “родовий”. За 3–5 діб до родів вульва набрякає і збільшується. За 2–3 доби густий слизовий корок шийки матки роз-
Розділ 10
ріджується і витікає за межі статевої щілини, пакети молочних залоз збільшуються і набрякають, в них з’являється молозиво.
За 15–40 годин до родів загальна температура тіла знижується на 0,4–1,2 ºС. Кролиця. За 1–3 доби до родів кролиця стає збудженою, бігає по клітці, висмикує
на животі і грудях свій пух і готує “гніздо для родів”. Апетит знижується, з’являються ознаки спраги. За 1–2 доби до родів пакети молочних залоз збільшуються і набряка-ють, в них з’являється молозиво.
Нутрія. Апетит нутрії за декілька днів до родів погіршується, живіт перед родами стає майже круглим; самки важко пересуваються, більше сидять у будиночку і тіль-ки на короткий час виходять купатися або для годівлі. Для нутрій гніздо утворюють штучно, закладаючи в будиночок сіно або солому, ущільнюючи підстилку руками до-верху, в якому нутрія робить прохід і в глибині влаштовує своє гніздо, яке захищає її новонароджених від холоду, особливо в зимову пору року. В основному роди у нутрії відбуваються вночі або дуже рідко вдень.
Сріблясто-чорна лисиця. За 1–2 доби до родів підвищується спрага. Поведінка самок перед родами буває різною: одні поводять себе спокійно, інші бігають з виском по клітці, шкребуться в будиночку. За 1–1,5 доби до родів самки вискубують пух на молочних залозах, а деякі обскубують увесь живіт і навіть хвіст. За 5–8 діб до щеніння у самок знижується апетит, що є ознакою нормальних (фізіологічних) родів. За 1–2 доби до родів молочні пакети у лисиць збільшуються і набрякають, в них з’являється молозиво. Іноді молозиво може з’являтися набагато раніше.
Дата добавления: 2016-01-04; просмотров: 43; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!