Ретровирусы (вирусы с обратной транскрипцией)
После проникновения происходит обратная транскрипция. Синтезированная ДНК – копия генома встраивается в геном клетки и использует клеточный механизм транскрипции

Параретровирусы
В капсиде в качестве генома содержится ДНК. ДНК сразу идет в ядро.Встраивается в геном клетки-хозяина с помощью интегразы.
ДНК - вирусы
Особенности жизненного цикла:
Ø Репликация и транскрипция происходит в ядре клетки (искл.: поксовирусы, асфавирусы).
Ø Для репликации вирусной ДНК необходима Ori – короткая нуклеотидная последовательность в вирусном геноме, к которым специфично присоединяются клеточные и вирусные факторы.
Ø Проблема недорепликации 5’-концов.

1)
2) Схема Кернса

3) Схема катящегося кольца
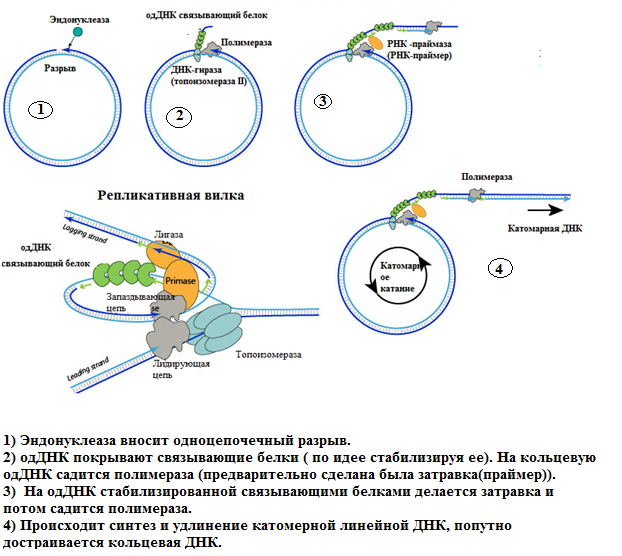
4) Самозатравочный механизм
Геном представлен линейной онДНК, оба конца которой имеют самокомплементарные (инвертированные повторы) последовательности, формирующие шпилечные структуры. Репликация вирусной ДНК осуществляется по самозатравочному механизму и осуществляется полимеразным комплексом из числа ранних вирусных белков: сначала эндонуклеаза вносит разрыв в цепи ДНК вблизи инвертированногоповтора с одного или двух концов (см. рис. 1.2.20, Б), после чего происходит расплетение концевой шпильки и синтез комплементарной цепи с использованием двунитевого участка в качестве затравки (см. рис. 1.2.20, В,Г), затем – отжиг концевых инвертированных повторов, которые используются как затравки для последующего синтеза комплементарных цепей ДНК (см. рис. 1.2.20, Д), в результате чего формируется либо пара геномов, либо конкатемер удвоенного вирусного генома (см. рис. 1.2.20, Е), и в последнем случае необходимо внесение двух эндонуклеазных разрывов цепи (см. рис. 1.2.20, Ж).

5) Аденовирусная репликация с использованием белков затравки

Особенностью репликации аденовирусной ДНК является её инициация без комплементарной олигонуклеотидной «затравки», а посредством белкового комплекса на 5'-конце, состоящем из ранних вирусных белков — ДНК-полимеразы (140 кДа), рТР (предшественника ТР) (75 кДа) и DBP (DNA-BindingProtein), а также клеточных: NFI (NuclearFactorI) и Oct-1 (Octamer-bindingprotein) (см. рис. 1.2.7, Б, Е). Синтез осуществляется непрерывно в направлении 5’ —> 3’ (см. рис. 1.2.7, В). Нереплицированная цепь ДНК (см. рис. 1.2.7) образует кольцо путём замыкания её комплементарных 5'- и З'-концов (см. рис. 1.2.7, Д), и двуцепочечный фрагмент используется для инициации синтеза ДНК (см. рис. 1.2.7, Е, Ж). Позже вирусная протеаза (23 кДа) осуществляет превращение рТР в ТР.

3. Тактические приёмы, используемые вирусами при экспрессии геномов. Сдвиг рамки считывания, протечка при сканировании, внутренняя инициация трансляции, рибосомальный пропуск, рибосомальное шунтирование.
Сдвиг рамки считывания – тип мутации в последовательности ДНК, для которого характерна вставка или делеция нуклеотидов, в количестве не кратном трем. В результате происходит сдвиг рамки считывания при транскрипции мРНК. Следует отличать от однонуклеотидного полиморфизма, в котором происходит замена одного нуклеотида на другой.
Протекающее сканирование –при сканировании мРНК большое значение имеет контекст AUG кодона (последовательность Шайна-Дельгана, Козак, разные трансляционные факторы). Если его ближайшее окружение не соответствует консенсусу, то с некоторой вероятностью (и эта вероятность тем больше, чем больше «несоответствие») сканирующий комплекс пропустит этот AUG и тогда синтез белковой цепи начнется со следующего AUG.
Рибосомальное шунтирование – Этот механизм инициации трансляции функционирует у нескольких вирусных мРНК. Трансляция этих мРНК осуществляется следующим образом: рибосомальная субъединица связывается с кэпом на 5'конце мРНК и начинает сканирование. 5'НТО таких РНК имеет выраженную вторичную структуру, но рибосомальная субъединица не «расплетает» ее, а каким-то образом обходит – шунтирует. Для успешного «шунтирования» вторичной структуры большое значение имеет «посадочная площадка» для рибосомы непосредственно перед AUG-кодоном. Однако самым важным является факт трансляции небольшой (состоящей всего из 3-х кодонов) рамки считывания, находящейся перед вторичной структурой. Таким образом, формально рибосомальное шунтирование является модификацией реинициации трансляции.
Внутренняя инициация трансляции (IRES) – регуляторная структура мРНК, задействованная в КЭП-независимом механизме инициации трансляции, при котором рибосома связывается с мРНК в области этой структуры в 5'-НТО недалеко от сайта инициации трансляции.
Рибосомальный пропуск – если после глицина идет пролин, происходит разрыв полипептида, при том что синтез полипептида продолжается. Вследствие этого, будут нарушены функции белка (или они изменятся) или же он станет нефункциональным.
Рибосомальный «пропуск»- это альтернативный механизм трансляции, в котором специфический вирусный белок мешает рибосоме от ковалентного связывания нового вставленного участка (см. 4), и дает возможность дальнейшей трансляции (см. 5). Это приводит к очевидному котрансплантационному расщеплению белка (см. 6).
Этот процесс вызывается «2А-like»/CHYSEL-последовательностью (cis-actinghydrolaseelement): Asp-Val/ Ile-Glu-X-Asn-Pro-Gly↓Pro (↓ - означает расщепление в этом месте).
Рибосомальный «пропуск» наблюдается только (+)ssRNA и dsRNA вирусов, чьи хозяева животные, растения или насекомые.

Дата добавления: 2018-06-01; просмотров: 1737; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!