Осевая мезодерма и боковая пластинка.
Осевая мезодермаподразделяется на сомиты и метамеризуется, причем закладка сомитов в разных классах хордовых неодинаков. У ланцетника закладываются жнетероцельно и сразу имеют целомическую полость, в то время как у большинства позвоночных сперва закладывается в виде сплошных скоплений мезодермальных клеток, и лишь потом появляются полости шизоцельным путем.
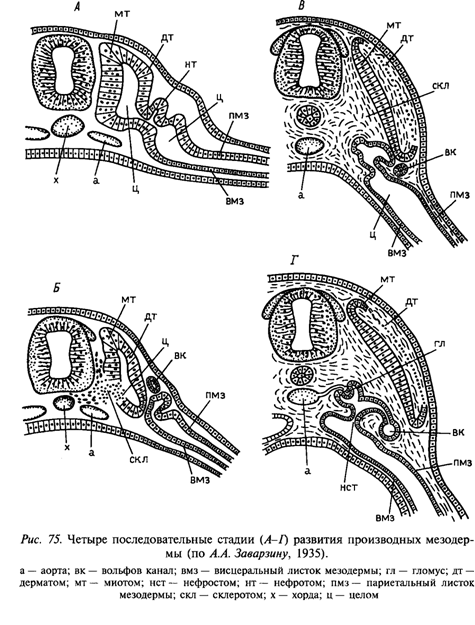
В ходе дальнейшего развития сомита из его клеток формируются три основные закладки:
- дерматом – наружная, обращенная к эктодерме. Формирует кожный листок, из него впоследствии дифференцируется соединительная часть кожи.
- склеротом – внутренняя часть сомита, примыкает к хорде (у низших позвоночных) или к хорде и нервной трубке (у высших). Зачаток осевого скелета, впоследствии распадающийся на отдельные клетки.
- миотом – располагается между дерматомом и склеротомом. Зачаток всей поперечно-полосатой мускулатуры.
У низших позвоночных основная часть сомитов – миотомы. У высших позвоночных наиболее активна зона, дающая как дерматом, так и миотом (он возникает путем подворачивания под зачаток дерматома).
Осевая мезодерма метамеризуется к в туловищном, так и головном отделах, но в последствии головные сомиты у позвоночных кроме ланцетника они распадаются, из их клеток формируются парахордалии, то есть по своей потенции они соответствуют склеротомам. Спереди от них возникают парные хрящевые закладки – трабекулы.
|
|
|
Часы сегментации.У зародышей костистых рыб, птиц и млекопитающих был обнаружен периодический процесс в несегментированной осевой мезодерме.
На примере куриного зародыша: спереди от гензеновского узелка в еще не сегментированной осевой мезодерме каждые 90 минут возникает область активации генов (один из них – c-hairy 1), начинается синтез мРНК. Область активации распространяется вперед, до границы уже сегментированной мезодермы, сзади ее догоняет область дезактивации, таким образом в конце от активной области остается лишь узкая полоса. Ровно в этот момент от несегментированной мезодермы отделяется новый сомит, и активная полоска остается в его составе.
Исследования показали, что клетки включают и выключают данную группу генов независимо от индукции, таким образом, часы действительно автономны.
Наличие часов сегментации не является необходимым условием для возникновения сомитов.

Развитие органов выделения.
У анамний два сменяющих друг друга органа выделения – головная почка (предпочка, пронефрос) и туловищная (первичная, мезонефрос).
У амниот вслед за ними развивается метанефрос (тазовая почка).
Все три типа образуются из мезодермы ножек сомитов:
- пронефрос из передних сомитов (наиболее четкая метамеризация; стенки канальцев непосредственно из ножек, поэтому канальцы открываются прямо в целом, имея на конце воронки-нефростомы, а другими концами сливаются в вольфов канал – первичный мочеточник; рост вольфова канала назад индуцирует образование мезонефрических канальцев)
|
|
|
- мезонефрос из почти всех туловищных сомитов (ножки к этому моменту отшнуровываются от сомитов и преобразуются в мезенхимную ткань; из них формируются метамерные канальцы с ответвлениями и изгибами; у высших позвоночных канальцы слепо заканчиваются в мезенхиме, а наружные концы открываются в первичный мочеточник; у высших позвоночных от них остаются только небольшие придатки половых желез)
- метанефрос из скопления нефрогенной мезенхимы, произошедшей из ножек сомитов (уже никаких следов метамерии; не связан с целомом ни на одной стадии развития; от первичного мочеточника в сторону нефрогенной мезенхимы идет остросток с расширенным концом, который превращается во вторичный мочеточник с почечной лоханкой)
Боковая пластинка.
Боковая пластинка располагается вентральнее ножек сомитов, рано разделяется на париетальный и висцеральный листки, соединенные брыжейками. Между ними находится целом, и оба листка формируют его выстилку – париетальный внешнюю, висцеральный внутреннуюю.
|
|
|
Производные висцерального листка(половые железы, сердце, кровеносные сосуды и клетки крови). Для полноценного развития нуждаются в контакте с энтодермой.
Половые железы и протоки.
Развиваются из висцерального листка боковой пластинки на уровне ножек сомитов – герминативного эпителия (половые клетки возникают из гоноцитов и мигрируют позже). Герминативный эпителий – соматическая ткань, формирующая стенки половой железы. Сама железа вначале представляет собой впячивание – половую складку, которая постепенно заполняется мезенхимой, из которой развивается мозговая часть железы.
Поначалу железа имеет одинаковое строение у обоих полов, но под воздействием проникших первичных половых клеток и гормонального баланса дифференцируется в семенник (преимущественно мозговой слой) или яичник (преимущественно корковая часть).
- У самцов семенные канальцы впадают в вольфовы каналы, которые становятся семяпроводами. У амниот это единственная функция вольфовых каналов (так как у тазовой почки вторичный мочеточник), у анамний они же являются мочеточниками.
|
|
|
- У самок имеются мюллеровы каналы (закладываются параллельно вольфовым, но у самцов дегенерируют), превращаются в яйцеводы.
У многих анамний развиваются верхние отделы мюллеровых каналов и открываются в целом воронкой нефростома, превратившейся в воронку яйцевода, так что яйцо сперва выходит в целом, а затем захватывается воронкой.
- Развитие сердца.
У птиц парный зачаток сердца возникает на вторые сутки инкубации в виде двух симметрично расположенных утолщений висцерального листка мезодермы, который тесно связан с энтодермой. Зачатки соединяются лишь после сворачивания энтобласта в трубку головной кишки. Из объединившейся висцеральной мезодермы возникает миокард. Эндокард формируется при слиянии трубчатых зачатков, образованных мигрировавшими мезенхимными клетками.
Единая сердечная трубка переходит в желточные вены, несущие кровь от стенки желточного мешка. Она лежит в перикардиальной полости, являющейся частью целома.
- Кровеносные сосуды. Развиваются из мезенхимы, закладываются в виде кровяных островков, отдельные трубочки сливаются в рыхлую сеть. Наружные клетки островков (ангиобласт) вступают в контакт друг с другом и формируют эндотелий, внутренние клетки (гемобласт) формируют клетки крови. Парные желточные вены впадают в зачаток сердца сзади и несут кровь от внезародышевых органов у амниот.
Ствол аорты делится на два артериальных ствола. Сначала формируется 6 дуг аорты по числу жаберных дуг, у высших позвоночных впоследствии большая часть редуцируется. Изначально формируется избыток сосудов, из которых потом остаются те, направление которых соответствует анатомическим особенностям тела взрослого животного.
Производные париетального листка –конечности.
- Конечности. Развиваются из выселившихся из париетального листка мезодермы мезенхимных клеток и покровной эктодермы.
У зародышей амфибий ранние зачатки конечностей имеют вид обособленных бугорков.
У зародышей амниот вначале формируются длинные складки (вольфовы гребни), которые позже редуцируются в средней части, а из переднего и заднего участка развиваются передние и задние конечности.
На ранних стадиях роста эктодермальный эпителий пассивно растягивается делящейся мезенхимой, но затем начинает активно участвовать в росте конечностей. У амниот эктодерма верхушки конечности утолщается, образуя апикальный гребешок.
Апикальная часть по мере роста расширяется и уплощается, зачаток скручивается вдоль своей продольной оси, на апикальной поверхности появляются зачатки пальцев (обособение пальцев связано с апоптозом клеток между ними).
Параллельно идет образование внутреннего скелета путем образования хрящей из мезенхимы. Выделяется стилоподия (зачаток проксимального хряща), их которой разовьются плечевые/бедренные кости; затем зигоподия (локтевой и лучевой хрящи; большой и малый берцовые) и аутоподия (хрящи кисти или стопы). Хрящи поясов конечностей формируются по времени между стило- и аутоподиями.
В конечность прорастают кровеносные сосуды и миобласты.
Интенсивные эпителио-мезодермальные взаимодействия.На начальной стадии мезодерма влияет на эктодермальный эпителий (он утолщается и начинает активно расти).
Нормальное развитие пальцев определяет обратное влияние на мезодерму со стороны эпителия верхушки почки конечности (апикального гребешка). При его удалении фаланги не дифференцируются. (При пересадке апикального гребешка на презумптивную мезодерму проксимальной части конечности развивается дистальный отдел; при пересадке проксимальной мезодермы задней конечности под гребешок передней, она даст дистальную часть задней конечности.)
Зона поляризующей активности– морфогенетически активная зона зачатка конечности на заднем крае, у основания. Если ее пересадить на передний край, то произойдет ее зеркальное удвоение – появится второй задний край с соответствующими пальцами. ЗПА характеризуется повышенной концентрацией морфогенетически активной ретиноевой кислоты (витамина А), градиент концентрации которой определяет передне-заднюю полярность.
В передних конечностях экспрессируется ген Tbx5, а в задних - Tbx4, продукты которых – факторы транскрипции, т.е. от них зависит активность других генов.
Важную роль играют белки группы факторов роста фибробластов. Для возникновения зоны поляризующей активности необходим ФРФ-10, а процессы апоптоза в пальцах контролируются белками ВМР.
Птицы.
Яйца птиц относятся к меробластическому типу, дробление дискоидальное. У верхнего полюса бластодиска формируется узкий клеточный слой – серп Коллара, который начинает расти в подзародышевую полость, в которую ранее выселяются отдельные клетки и образуют первичный гипобласт. Затем первичный гипобласт объединяется с серпом Коллара во вторичный гипобласт(носитель передне-задней полярности зародыша). Гипобласт дает начало внезародышевой энтодерме, которая обрастает желток и участвует в формировании желточного мешка.
Наружный слой бластодиска после выселения из него клеток называется эпибластом.
 Центральная зона бластодермы после разделения выглядит прозрачной и носит название area pellucida. Перибласт (краевое кольцо бластодермы) выглядит темным и называется area opaca. На заднем конце зародыша между этими двуми зонами возникает первичная полоска, после чего приобретает углубление и начинает называться первичной бороздкой. На переднем конце возникает первичная ямка, или гензеновский узелок (из области узелка начинают иммигрировать под эпибласт клетки дефинитивной кишечной энтодермы - энтобласта).
Центральная зона бластодермы после разделения выглядит прозрачной и носит название area pellucida. Перибласт (краевое кольцо бластодермы) выглядит темным и называется area opaca. На заднем конце зародыша между этими двуми зонами возникает первичная полоска, после чего приобретает углубление и начинает называться первичной бороздкой. На переднем конце возникает первичная ямка, или гензеновский узелок (из области узелка начинают иммигрировать под эпибласт клетки дефинитивной кишечной энтодермы - энтобласта).
Формирование энтобласта – начало второй стадии гаструляции. Клетки энтобласта оттесняют гипобласт на периферию зародышевого щитка. Вслед за энтобластом через гензеновский узелок и стенки первичной бороздки мигрируют клетки будущего мезобласта (среднего зародышевого листка). Эпителио-иезенхимный переход при миграции.
Центральная часть мезобласта, мигрировавшая через гензеновский узелок дает начало головной мезодерме и хордальному выросту (и затем хорде).
Материал, мигрировавший через боковые стенки первичной бороздки, дает начало внезародышевой мезодерме (кровяные островки и сосуды).
Материал, мигрирующий последним (ближе к оси зародыша) даже мезодерму боковой пластинки, а самый поздний – сомиты.
Бороздка постепенно укорачивается, узелок смещается назад, а затем превращается в анальное отверстие.
Вторая фаза гаструляции гомологична гаструляции амфибий. Первичная бороздка гомологична бластопору, ее края – губам бластопора, гензеновский узелок – дорсальной губе бластопора (подтврждается экспериментально; первичные организаторы гаструляции). Материал энтобласта гомологичен прехордальной пластинке амфибий.
Дата добавления: 2018-05-09; просмотров: 1633; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!