Оплодотворение. Молекулярные процессы при активации яйцеклетки.
Активация яйцеклеток - кортикальная реакция. Ее биохимические основы.
1) Моментом начала активации яйцеклетки считается образование бугорка оплодотворения.
Происходит благодаря полимеризации кортикального актина. Бугорок оплодотворения формируется после рецепции биндинов рецепторами желточной оболочки яйца.
2) Мембраны сперматозоида и яйцеклетки сливаются, образуется канал, по которому затем пройдут ядро и центриоль сперматозоида. В мембране сперматозоида, оказавшейся частью этого канала, находятся Na+ каналы, по которым натрий поступает в цитоплазму яйца. Мембранный потенциал яйцеклетки на несколько секунд меняется с -70 мВ на + 10 мВ => быстрый блок полиспермии. Это объясняется тем, что положительно заряженная мембрана яйцеклетки для сперматозоидов непроницаема (наличие потенциал-зависимых рецепторов).
3) При контакте биндина с мембранным рецептором яйцеклетки меняется конформация G-белка. Он активирует фосфолипазу С, которая также встроена в мембрану. Она расщепляет фосфатидил-инозитол-4,5-дифосфат(PIP2) на диацилглицерол, DAG, (он затем через протеинкиназу С стимулирует транспорт Na+ внутрь клетки, H+ - наружу, что приводит к подъему синтеза белка в цитоплазме яйцеклетки и синтез ДНК в мужском и женском ядрах) и инозитол-3-фосфат (IP3). Последний стимулирует выход Ca2+ из ЭПР. Кальций стимулирует экзоцитоз кортикальных гранул (альвеол) яйцеклетки (происходит кортикальная реакция). При этом в пространство между плазмалеммой и желточной оболочкой высвобождается ряд веществ:
|
|
|
1) вителлиновая деламиназа. Отделяет желточную облочку от плазмалеммы путем лизирования динеина, их соединяющего.
2) Спермрецепторная гидролаза лизирует сайты соединения налипших сперматозоидов, очищая от них поверхность яйца.
3) Осмотически активный гликопротеид, приводяший к поступлению воды в щель между плазмалеммой и желточной оболочкой => образованию перивителлинового пространства.
4) Фактор, способствующий затвердеванию желточной оболочки (теперь она называется оболочкой оплодотворения и становится недоступной для сперматозоидов). Медленный блок полиспермии.
5) Гиалин – структурный белок, участвующий в формировании гиалинового слоя, располагающегося у яйцеклеток некоторых видов над плазмалеммой.
Помимо этого, увеличение концентрации Ca2+ приводит к:
• активации синтеза белка (особенно гистонов и циклинов), ДНК (совместно с DAG)
• сборке и перераспределение элементов цитоскелета, в результате чего кортикальный слой приобретает сократимость, необходимую для дробления
• активация NAD-киназы (путем изменения активности глюкозо-6-фосфатдегидрогеназы) => образование NADP => стабилизация оболочки оплодотворения (за счет синтеза липидов)
|
|
|
• активация кальпина II.
Фаза зрелости яйцеклеток различных групп животных при проникновении сперматозоида.
Мейоз завершается еще во время оогенеза у морских ежей и некоторых кишечнополостных.
У остальных видов происходит блок мейоза, который снимается только с оплодотворением.
1) Диакинез профазы I. Губки, плоские, круглые, кольчатые черви, моллюски, щетинкочелюстные, лиса, собака и лошадь.
2) Метафаза I - губки, немертины, кольчатые черви, моллюски, почти все насекомые.
3) Метафаза II – почти все хордовые. (У летучих мышей – анафаза II).
В состоянии метафазы II яйцеклетку удерживает белковый комплекс MPF, деградацию которого предотвращает циклин В. А его защищает от деградации pp39mos.
Ооплазматическая сегрегация при дроблении.
После проникновения сперматозоида начинаются процессы расслоения, отмешивания различных составных частей цитоплазмы яйцеклетки (ооплазмы) – ооплазматическая сегрегация.
Намечается радиальная (относительно анимально-вегетативной оси) симметрия.
У многих беспозвоночных разные бластомеры дают начало строго определенным зачаткам.
|
|
|
Видимо, это обусловлено ооплазматической сегрегацией при дроблении и взаимодействиями между бластомерами.
Движения цитоплазмы хорошо заметны в тех случаях, когда разные участки цитоплазмы содержат разноцветные гранулы (желток, темный пигмент и др.). Например, у гребневиков по периферии яйцеклетки располагается зеленая цитоплазма (до стадии 8 бластомеров). После деления становятся неравномерными: крупные бластомеры отпочковывают от себя микромеры, которые целиком состоят из зеленой эктоплазмы. Микро- и макромеры резко отличаются по своей будущей судьбе и не способны ее изменить.
При спиральном дроблении (например, кольчатые черви и моллюски) на вегетативном полюсе есть полярная плазма. После 3 деления она вся попадает в один бластомер (1D, вегетативное полушарие, спинная сторона). Позже она большей частью попадает в 2d и 4d, из которых развивается большая часть эктодермы и целомическая мезодерма. Те, при удалении полярной плазмы, возникают личинки, лишенные мезодермы и некоторых эктодермальных закладок.

9. Дробление. Типы и закономерности пространственного расположения бластомеров.
Дробление начинается после оплодотворения и активации. Дробление – ряд непрерывно следующих друг за другом митотических делений, но рост отсутствует. Образуется полость внутри – бластоцель.
|
|
|
Положение частей цитоплазмы не меняется, но они попадают в разные бластомеры. Ядерно-плазменное отношение в начале дробления достигает уровня, характерного для обычных соматических клеток.
Дробление(синхронность и асинхронность указывают на перестройку клеточных циклов и отражают синтетическую активность бластомеров):
- синхронное (ранние стадии, первые 4-7 раз, число бластомеров 2^n)
- асинхронное (сначала идет волнами; пробуждается транскрипционный аппарат зародыша, появляется фаза G1; иногда эту фазу называют бластуляцией)
Функции дробления:
- формирование многоклеточности
- увеличение ядерно-плазменного отношения (которое сильно уменьшилось во время большого роста)
Особенности клеточного цикла:
- синтез ДНК начинается уже в пронуклеусах (все факторы инициации синтеза и репликации (полимеразы, гистоны и др) уже есть в яйце с оогенеза), начинается еще в телофазах.
- Клеточные циклы укорочены за счет G1 и G2 периодов (опять же потому, что все нужные продукты есть в яйце).
- S период укорочен за счет полирепликонности клеток эукариот – каждый репликон может реплицироваться автономно (большая полирепликонность, чем у обычных эукариотических клеток).
Цитотомия- осуществляется с помощью двух механизмов:
- образование кольца из сократимых микрофиламентов (образуется в кортикальном слое клетки, в плоскости, перпендикулярной прямой, соединяющей клеточные центры; начинает деление цитоплазмы; ярче выражен у маложелтковых клеток)
- встраивание запасенных предшественников цитоплазмы (сильнее у многожелтковых клеток)
Регуляция дробления. Фактор созревания (пик концентрации в митозе, самая низкая – в S-фазе), который регулируется цитостатическим фактором (стабилизирует фактор созревания, задерживая клетки в митозе), а тот ингибируется ионами Са.
Пространственная организация дробления.Определяется:
- закономерным расположением интерфазных ядер
- закономерной ориентацией веретен деления
- движениями бластомеров
Закон Гертвига-Сакса:
- клеточное ядро стремится расположиться в центре чистой, свободной от желтка цитоплазмы.
- веретено клеточного деления стремится расположиться по направлению наибольшего протяжения свободной от желтка цитоплазмы.
Принято считать, что характер дробления зависит от типа яйцеклетки.
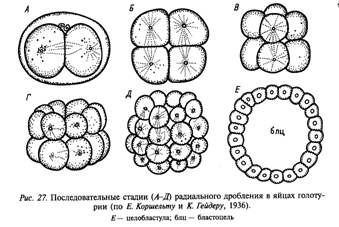
Дробление:
1. Полное (голобластическое)
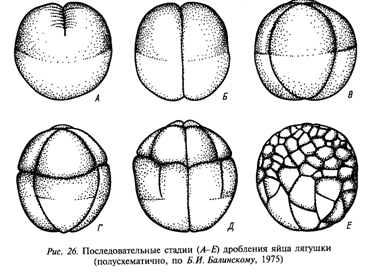
1.1. Равномерное (для олиго- и изолецитальных клеток). Объемы бластомеров одинаковы. Целобластула – однослойная бластодерма с крупным бластоцелем в центре.
 1.2. Неравномерное (для мезо- и тело). Объемы бластомеров неодинаковы. Амфибластула – состоит из микро- и макромеров, бластоцель сдвинут к анимальному полюсу (к микромерам).
1.2. Неравномерное (для мезо- и тело). Объемы бластомеров неодинаковы. Амфибластула – состоит из микро- и макромеров, бластоцель сдвинут к анимальному полюсу (к микромерам).
2. Неполное (меробластическое)
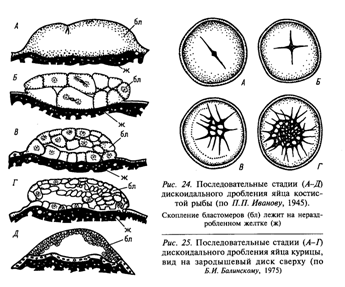
2.1. Дискоидальное (поли- и тело-). Борозды формируются вслед за делениями ядер, но разделяют только один из полюсов яйца. Дискобластула – диск бластомеров, лежащий на желтке.
 2.2. Поверхностное (поли- и центро-). Поверхностный слой цитоплазмы разделяется перегородками, перпендикулярными поверхности яйца, центральная часть не делится. Перибластула – не имеет бластоцеля – внутри желток.
2.2. Поверхностное (поли- и центро-). Поверхностный слой цитоплазмы разделяется перегородками, перпендикулярными поверхности яйца, центральная часть не делится. Перибластула – не имеет бластоцеля – внутри желток.
Расположение образующихся из зиготы бластомеров в пространстве друг относительно друга послужило основанием для определения типов дробления, исходя из расположения бластомеров. Выделяют радиальное (ланцетник), спиральное (моллюски), билатеральное, или двусторонне-симметричное (круглые черви), бисимметричное, или двусимметричное (гребневики), и анархичное (плоские черви) дробления.
При радиальном дроблении борозды деления (митотические веретена) ориентированы параллельно или перпендикулярно анимально-вегетативной оси яйцеклетки. Через такую бластулу проходит несколько плоскостей (осей) симметрии.
Спиральное дробление отличается нарушением такого соответствия (борозды деления располагаются наклонно к анимально-вегетативной оси), и дочерние бластомеры располагаются как бы по спирали. Образующаяся при спиральном дроблении бластула (стереобластула) не имеет ни полости, ни даже одной плоскости симметрии.
Билатеральное дробление характеризуется наличием в формирующейся бластуле одной оси (плоскости) симметрии. При бисимметричном дроблении формирующаяся бластула имеет две оси (плоскости) симметрии.
Анархичное деление резко выделяется от описанных выше неупорядоченным расположением бластомеров и отсутствием оси (плоскости) симметрии.
Дата добавления: 2018-05-09; просмотров: 805; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!