Молекулярные механизмы первичной эмбриональной индукции. Индукция по умолчанию.
Эмбриональная ииндукция – целый каскад генов, активирующий или ингибирующий друг друга, и шпемановская и ньюкуповская индукция – части такого каскада.
Важные вещества: гликопротеиды семейства Wnt-1 и надсемейство ФРО-бета (фактор роста опухолей). Белок Vg1 вызывает ньюкуповскую индукцию мезодермы.
Белок бета-катенин, распределенный в цитоплазме зиготы более-менее равномерно, после оплодотворения подвергается ферментативному расщеплению, но на дорсальной стороне расщепление подавлено другим белком. Бета-катенин сохраняется и попадает в ядра бластомеров, становясь фактором ньюкуповской индукции. Он связывается с промоторами опредленных генов и активирует их, например, определяя лево-правую асимметрию. Другой активируемый ген производит белок, соединяющийся с ФРО-бета и активирует ядерный белок goosecoid, он содержится в ядрах клеток шпемановского индукционного центра, активируя работу других генов, являющихся непосредственными факторами шпемановской индукции.
Белки семейства ВМР, входящие в то же надсемейство, концентрируются на вентральной стороне зародыша, секретируются в межклеточное пространство и, связываясь с рецепторами, запрещают клеткам дифференцироваться в нервную ткань и другие производные осевой мезодермы.
Индукция «по умолчанию» - дифференцировка в сторону нейральных производных. Если не препятствовать связыванию белков группы Wnt с клетками презумптивной нейральной эктодермы, то вся нервная пластинка развивается в спинной мозг. Для формирования головного мозга необходимо связать факторы Wnt в межклеточном постранстве, что и осуществляется головными индукторами — белками Cerebrus и Dickkopf, секретируемыми прехордальной пластинкой. Если такое связывание произошло, то в области будущего головного мозга активируется ген OTX-2. Однако у всех позвоночных на последующих стадиях развития (ранняя и средняя нейрула) активность этого гена блокируется и активизируется «ген переднего конца» - anf. Тогда осуществляется нормальное развитие головного мозга.
|
|
|
При первичной эмбриональной индукции индуктор "приказывает" индуцируемой ткани, в каком направлении ей изменяться. Подобные взаимодействия были названы инструктивными. При инструктивных взаимодействиях реагирующая ткань под воздействием индуктора развивается в строго определенном направлении. В отсутствии индуктора развития в этом направлении не происходит. При пересадке другой ткани на место индуктора развития в данном направлении тоже нет. Наконец, под влиянием индуктора другие ткани могут развиваться в том направлении, которое задается индукцией. Взаимодействия такого типа играют важную роль и на более поздних стадиях развития. Они были названы вторичной индукцией.
|
|
|
Роль альтернативного сплайсинга в регуляции клеточной дифференцировки. Регуляция на уровне трансляции. Роль микроскопических РНК.
Регуляция на уровне сплайсинга может происходить еще во время транскрипции.
Альтернативный сплайсинг - по-разному сшитые первичные транскрипты одного гена дают разные мРНК. Те, в свою очередь, могут играть роль в разных дифференцировках: например, для первичного транскрипта альфа-тропомиозина мыши путем различных сшивок от него можно получить мРНК для скелетных мышц, для клеток мозга и для фибробластов.
Ген дрозофили DSCAM альтернативным сплайсингом дает 38 000 белков.
Альтернативный сплайсиг контролируется макромолекулярным комплексом -- сплайсосомой -- состоящей из белков и малых РНК.
Некоторые ученые считают данный уровень регуляции одним из самых важных.
Регуляция на уровне трансляции: проявляется в основном в задержке трансляции заготовленных ранее мРНК. Например, блок трансляции заготовленных в оогенезе мРНК вплоть до активации яйцеклетки. Устранение блока трансляции после активации яйцеклетки достигается добавлением большого числа адениловых групп на 3'-конце молекулы мРНК.в ходе дробления мРНК вступает в работу не сразу, а по определенной программе, регулируется разнообразными факторами (у моллюсков, например, он связан с полярной лопастью).
|
|
|
Существенные задержки в транскрипции отмечаются также у таких специализированных клеток - эритроидных, сперматогенных и др. Кроме этого, уровень экспрессии на разных стадиях дифференцировки сильно разнится.
Регуляция на уровне трансляции наблюдается также у гемоглобина в силу сложности его строения (2 альфа-, 2 бета-глобиновых цепей и 4 молекулы гема). Любое отклонение фатально.
Избыток гема блокирует ключевой фермент, который отвечает за его синтез. Этот же избыток активирует синтез глобинов на уровне трансляции т.к. ингибирует протеинкиназу (та, в свою очередь, инактивирует фосфорилированием фактор инициации трансляции глобина). Ингибирование ингибитора приводит к активации. Таким образом поддерживается нормальное соотношение компонентов гемоглобина.
Еще один аспект сложной регуляции синтеза гемоглобина заключается в том, что в диплоидной клетке есть 4 активных альфа-глобиновых и только 2 активных бета-глобиновых гема, а соотношение концентации самих белков поддержиается на уровне 1:1. Эта регуляция осуществляется на уровне трансляции за счет конкуренции обеих мРНК за факторы инициации трансляции.
|
|
|
Еще один способ регуляции на уровне трансляции - РНК-интерференция, действующая в ходе развития круглого червя Caenorhabditis elegans (аналогичный механизм найден в эмбриональных стволовых клетках мышей). Для нормально развития этого червя необходимо подавление деления стволовых клеток. Оно происходит благодаря активности определенных генов и последующему сплайсингу (возникают короткие молекулы РНК, которые комплементарны тем молекулам мРНК, которые вовлечены в трансляцию). В итоге эти РНК связывают мРНК, и трансляция подавляется. Иногда перед этим мРНК деградируют.
Гаструляция у амфибий.

Предгаструляионные движения.Вегетативная стенка сложена богатыми желтком макромерами, поэтому впячивание незначительно. Такие иммиграционные движения приводят к сокращению светлой вегетативной зоны и увеличению темной анимальной. Можно рассматривать как первую фазу эпиболии.

Собственно гаструляция.Начинается в области серого серпа. Образуется линия выровненных клеточных стенок, а затем по этой линии формируется узкая щель – зачаток бластопора. Она углубляется, внутрь втягиваются все новые клетки поверхности, принимает вид серповидной бороздки. Анимальная область становится спинной (дорсальной) губой. Полость щелевидной бороздки расширяется и превращается в полость первичной кишки – архентерона (гастроцель).
Подворачивание клеточного материала через дорсальную губу.Клетки анимальных районов перемещаются вегетопетально до губы бластопора, а затем подворачиваются через нее и формируют выстилку растущего архентерона. Является продолжением эпиболических процессов.
Эпиболия.Основное движение эпиболии – интеркаляция клеток (переупаковка с утратой старых межклеточных контактов и образованием новых):
Радиальная интеркаляция - протекает с самого начала гаструляции вдоль крыши бластоцеля; изначально у бесхвостых крыша из 3-4 слоев клеток, но в ходе радиальной интеркаляции все слои кроме наружного смещаются радиально во встречных направлениях; крыша истончается, но ее общая площадь растет; у хвостатых крыша однослойная, так что клетки уплощаются и истончаются, что приводит к тому же результату.
Планарная конвергентная интеркалаяция –с начала видимой гаструляции, происходит в супрабластопоральной области. Клетки движутся во встречных направлениях к дорсомедиальной линии, где интердигитируют. Происходит растяжение супрабластопоральной области, которая становится спинной стороной.
Бластопор продолжает рост. Он охватывает вегетативную зону сперва полукольцом, а затем и кольцом, которое постепенно уменьшается до узкого отверстия. Заключенный внутри бластопора светлый вегетативный материал называется желточной пробкой. Выделяются вентральная и боковые губы, через них тоже идет подворачивание, но более слабое.
Бластоцель оттесняется продвигающейся внутрь стенкой архентерона в вентральном направлении. Это движение инвагинации, оно слагается из следующих компонентов:
- закладка бластопора –возникновение группы колбовидных клеток с узкими апикальными шейками. Клетки способны менять форму, благодаря этому и происходит закладка.
- клетки вершины гастрального впячивания активно ползут по стенке бластоцеля, на которой формируются направляющие волокна внеклеточного матрикса из фибронектина (по завершении инвагинации колбовидные клетки уплощаются и образуют выстилку передней части эмбрионального кишечника, а в будущем - печень).
- клетки внутренних слоев губы бластопора при подворачивании из столбчатых эпителиальных превращаются в округлые разобщенные. Клеточный слой рассыпается (удобно для подворачивания)
- смещение губы в вегетативном направлении (эпиболия быстрее подворачивания)
- удлинение деламинационной борозды, разграничивающей вернувшийся и еще не вернувшийся клеточный материал
У бесхвостых.Колбовидные клетки инициируют начало движения, а затем могут быть удалены.
У хвостатых.Колбовидные клетки играют ведущую роль в миграции.

34. Внезародышевые органы млекопитающих и типы плацент.
У плацентарных млекопитающих яйца алецитальные, дробление полное, но хрен знает какое (нельзя отнести ни к одному типу). Дробление с самого начала асинхронное, геном зародыша начинает функционировать очень рано. Образуется стерробластула. В стерробластуле оченб скоро выделяется слой светлых наружных клеток и темный – внутренних в виде плотной массы (зародышевого узелка).
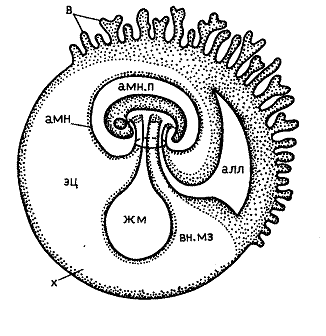 Из наружного слоя впоследствии развивается внезародышевая ткань – трофобласт. Полость амниона в большинстве случаев возникает кавитационным (шизоцельным путем) благодаря расхождению клеток.
Из наружного слоя впоследствии развивается внезародышевая ткань – трофобласт. Полость амниона в большинстве случаев возникает кавитационным (шизоцельным путем) благодаря расхождению клеток.
Первичная полоска дает начало внезародышевой мезодерме. Во внезародышевой мезодерма развиваются лакуны, которые, сливаясь, образуют полость внезародышевого целома. В трофобласте развиваются первичные ворсинки,в которые затем врастают клетки внезародышевой мезодермы, образуя кровеносные сосуды (и они превращаются во вторичные ворсинки). Трофобласт со вторичными ворсинками называется хорионом.
Позже развивается аллантоиндная ножка из внезародышевой мезодермы. Богата кровеносными сосудами, подрастает изнутри к ворсинкам хориона (относится к зародышевой части плаценты, особенно важна для сумчатых, где материнская часть плаценты неразвита).
Имплантация – погружение вторичных ворсинок хориона в стенку матки.
Продолжением имплантации является процесс образования плаценты – плацентация.
Плацента – провизорный орган, который формируется во время беременности, в ее построении участвуют как эмбриональные, так и материнские ткани. Через плаценту осуществляется связь зародыша с материнским организмом.
Функции плаценты:
- трофическая
- газообмен
- защитная
- регуляторная
- гормональная
- антитоксическая и т.д.
Основная функция плаценты заключается в передаче веществ, растворенных в крови матери, эмбриону и наоборот. Кроме того, плаценты многих млекопитающих вырабатывают гормоны, которые способствуют сохранению беременности (у человека это хорионический гонадотропин). При рождении плода плацента отторгается.
Классификации плацент:
По глубине погружения вторичных ворсинок:
- полуплацента(свинья, лошадь, китообразные) – ворсинки не прободают даже эапителиальной выстилки матки, только погружаются в складки ее слизистой оболочки.
- десмохориальная(у жвачных) – ворсинки разрушают слизистую и внедряются в соединительнотканный слой, но не достает до стенок сосудов матки
- эндотелиохориальная(у хищных) – установление контакта между сосудами плода и матери, отделяются от ворсинок только эндотелием
- гемохориальная(приматы) – погружение ворсинок в кровяные лакуны с кровью матери, таким образом, кровь отделяется от крови матери только оболочкой хориона и стенкой капилляров плода. Клетки ворсинок хориона способны к пиноцитозу. Имеется плацентарный барьер, препятствующий проникновению некоторых веществ.
Классификация по О.Гертвигу:
- Диффузная плацента - характерно образование ворсин по всей поверхности хориона
- Множественные плаценты – хориальные ворсины представлены группами, границами между ними являются участки гладкого хориона
- Зонарная плацента– ворсины хориона располагаются, как бы опоясывая плод
- Дискоидальная плацента – хориальные ворсины сконцентрированы в одном участке хориона; характерны для грызунов, приматов и человека
По степени повреждения материнских тканей при родах все типы плацент подразделяют на отпадающие (или децидуальные) и неотпадающие.
Для свиней, лошадей, верблюдов, многих жвачных животных, лемуров и некоторых других видов плацентарных млекопитающих характерна неотпадающая плацента, состоящая только из плодных тканей. При родах ворсины хориона выходят из углублений слизистой оболочки матки, не повреждая ее, без кровотечений.
Для хищных, грызунов, насекомоядных, рукокрылых, приматов и человека характерны плаценты отпадающего (децидуального типа). В состав такой плаценты входят как плодные, так и материнские ткани. При родах происходит так называемое отторжение плаценты – вместе с ворсинами хориона отторгается и часть слизистой оболочки матки, что вызывает довольно значительное кровотечение. 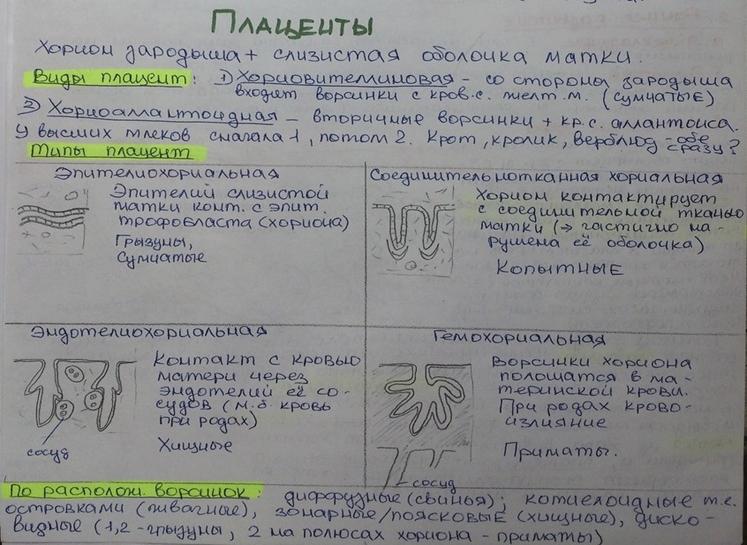
Дата добавления: 2018-05-09; просмотров: 890; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!