Особенности развития центральной нервной системы
Нервная система начинает развиваться на 3-й неделе внутриутробного развития из эктодермы (наружного зародышевого листка).
На дорсальной (спинной) стороне зародыша исходит утолщение эктодермы. Это формируется нервная пластинка. Затем нервная пластинка изгибается вглубь зародыша и образуется нервная бороздка. Края нервной бороздки смыкаются, формируя нервную трубку. Длинная полая нервная трубка, лежащая сначала на поверхности эктодермы, отделяется от нее и погружается внутрь, под эктодерму. Нервная трубка расширяется на переднем конце, из которого позднее формируется головной мозг. Остальная часть нервной трубки преобразуется в спинной мозг (рис. ) .
Из клеток, мигрирующих из боковых стенок нервной трубки, закладываются два нервных гребня- нервные тяжи. В дальнейшем из нервных тяжей образуются спинальные и автономные ганглии и шванновские клетки, которые формируют миелиновые оболочки нервных волокон. Кроме того, клетки нервного гребня участвуют в образовании мягкой и паутинной оболочек мозга. Во внутреннем слое нервной трубки происходит усиленное деление клеток. Эти клетки дифференцируются на два типа: нейробласты (предшественники нейронов) и спонгиобласты (предшественники глиальных клеток). Одновременно с делением клеток головной конец нервной трубки подразделяется на три отдела - первичные мозговые пузыри. Соответственно они называются передний (I пузырь), средний (II пузырь) и задний (III пузырь) мозг. Передний мозговой пузырь является родоначальником больших полушарий головного мозга. Средний мозговой пузырь формирует подкорковые образования, включая такие крупные, как таламус, гипоталамус, а также средний мозг, который представляет собой ножки мозга, соединяющие большие полушария со стволом. Задний мозговой пузырь является предшественником ствола головного и спинного мозга.
К концу 4-й недели стенка нервной трубки состоит из трех слоев: наружного, мантийного (среднего) и эпендимного (внутреннего). Нарастание клеточной массы в нервной трубке происходит за счет внутреннего эпендимного слоя, содержащего нейробласты и спонгиобласты. Размножение и созревание вновь образованных нейробластов и глиоцитов происходит в мантийном слое, а в тонком наружном слое располагаются волокна и верхушечные дендриты созревающих нейронов.
Нервные узлы черепных нервов, так же, как и сенсорные узлы спинномозговых нервов, образуются из нервного гребня - длинного тяжа, расположенного по обеим сторонам крыльных пластинок нервной трубки и состоящего из клеток нейроэктодермы. Нервный гребень, доходящий в краниальном отделе нервной трубки до среднего мозга, дает начало периферическим постганглионарным нейронам ресничного, крылонебного, подчелюстного и подъязычного вегетативных узлов и их нервным сплетениям, а также сенсорным нейронам в составе узлов сенсорного (VIII пара) и бранхиогенных (V, VII, IX, X пары) черепных нервов.
Соматомоторные (III, IV, VI, XII пары) черепные нервы, а также двигательные волокна в составе бранхиогенных черепных нервов (V, VII, IX, X, XI пары), иннервирующих мышцы жаберного происхождения, формируются аксонами нейронов, развивающихся из клеток базальной пластинки среднего и заднего мозговых пузырей. Парасимпатические преганглионарные нейроны вегетативных ядер III, VII, IX и X пар черепных нервов развиваются из межуточной зоны, расположенной между крыльной и базальной частями стенок среднего и заднего мозговых пузырей.
К концу 3-й недели эмбриогенеза, еще до завершения нейруляции и формирования мозговых пузырей в результате быстрого роста краниального отдела нервной трубки происходит ее изгиб вперед и вверх. Затем формируются и другие изгибы нервной трубки.
В последующем развитии передний мозг делится на конечный (большие полушария) и промежуточный мозг. Средний мозг сохраняется как единое целое, а задний мозг делится на два отдела, включающих мозжечок с мостом и продолговатый мозг. Это 5-пузырная стадия развития мозга.
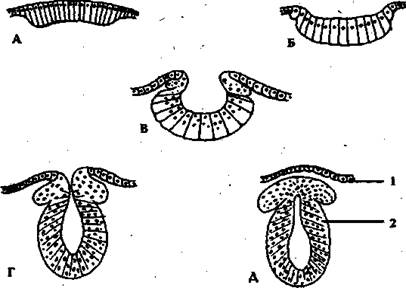
Рис. . Стадии эмбриогенеза нервной системы в поперечном схематическом разрезе:
a - медулярная пластина; б, г - медулярная бороздка; в, д - мозговая трубка;
1 - роговой листок (эпидермис); 2- ганглиозный валик.
Схема развития головного мозга
| Наименование отделов | Трехпузырная стадия | Пятипузырная стадия | Наименование отделов | Структуры, составляющие данные отделы | |
| Передний мозг | 1-й мозговой пузырь (передний) | 1-й мозговой пузырь | Конечный мозг | Большие полушария головного мозга | Кора больших полушарий, белое вещество больших полушарий, базальные ганглии, боковые желудочки |
| 2-й мозговой пузырь | Промежуточный мозг | Зрительные бугры, надбугорная область, забугорная область, подбугорная область | |||
| Глазные пузыри | Глазные бокалы, сетчатка глаза, зрительный нерв | ||||
| Средний мозг | 2-й мозговой пузырь (средний) | 3-й мозговой пузырь | Средний мозг | Ножки мозга (из вентральной части), четверохолмия (из дорсальной части) | |
| Ромбовидный мозг | 3-й мозговой пузырь (задний или ромбовидный) | 4-й мозговой пузырь | Задний мозг | Верхние ножки мозжечка (из передней части), мост (из вентральной части), мозжечок (из дорсальной части) | |
| 5-й мозговой пузырь | Продолговатый мозг | Продолговатый мозг | |||
Последующее развитие мозговых пузырей имеет свои особенности. В начале 5-й недели продолговатый мозг почти не отличается от спинномозгового отдела нервной трубки: он имеет крыльную и базальную пластинки. В толще базальной и крыльной пластинок происходит закладка ядер V-XII пар черепных нервов, причем моторные ядра формируются из материала базальной пластинки, поэтому располагаются медиальнее по отношению к сенсорным ядрам - производным крыльной пластинки. Формирующиеся вегетативные ядра черепных нервов занимают промежу-
Таблица .
Развитие отделов головного мозга
| Первичный отдел | Вторичный отдел | Полости мозга | Нервы | Области иннервации |
| Нервная трубка | Спинной мозг | Центральный канал | Спинномозговые (31 пара) | Туловище: кожный покров, мышцы, внутренние органы |
| Ромбовидный мозг | Продолговатый мозг | IV желудочек | Черепные XII ,X, XI, IX пары | Глотка, язык и его мышцы , гортань, другие органы |
|
Задний мозг Мост
Мозжечок
| VIII, VII, VI, V пары | Кожный покров лица и головы, жевательные мышцы, мимические мышцы, орган слуха | ||
| - | - | |||
| Средний мозг | Средний мозг | Водопровод мозга | III, IV пары | Мышцы глазного яблока |
| Передний мозг | Промежуточный мозг | III желудочек | II пара | Орган зрения |
| Конечный мозг | Боковые желудочки | I пара | Орган обоняния |
точное положение между моторными и сенсорными. Одновременно с закладкой ядер, а также дифференцировкой клеток нервного гребня происходит развитие самих черепных нервов.
В развитии заднего мозга прослеживаются те же изменения в расположении пластинок нервной трубки, что и в продолговатом мозге. Вследствие расхождения крыльных пластинок в стороны здесь также происходит истончение крыши нервной трубки.
Миграция клеток крыльной пластинки в область крыши заднего мозга приводит к формированию на 12-16-й неделях червя и полушарий мозжечка. Под мозжечком просматривается полость будущего IV желудочка, дно которой образует ромбовидная ямка.
Средний мозг обособляется относительно рано, на 4-й неделе развития. Клетки крыльной пластинки в области среднего мозга мигрируют в дорсальном и вентральном направлениях. На дорсальной поверхности они дают начало ядрам верхних и нижних холмиков пластинки будущего четверохолмия. Клетки, смещающиеся вентрально, формируют красные ядра и ретикулярные ядра среднего мозга. Из базальной пластинки образуется также ядро
глазодвигательного нерва (III пара), хорошо выраженное на 12-й неделе внутриутробного развития.
Промежуточный мозг обособляется от переднего мозгового пузыря на 5-й неделе. В ростральной (передней) части нервной трубки, дающей начало конечному и промежуточному мозгу, идет интенсивное увеличение числа нервных клеток. Здесь отсутствует деление на крыльную и базальную пластинки, не происходит формирования черепных нервов, а вся масса нервных клеток идет на развитие многочисленных ядер и коры головного мозга.
В промежуточном мозге на 5-й неделе наблюдается начало интенсивного роста мантийного (пролиферативного) слоя, что приводит к значительному утолщению его латеральных стенок; здесь образуются закладки правого и левого таламусов. Еще до обособления промежуточного мозга в задней части переднего мозгового пузыря на его вентролатеральной поверхности появляются парные глазные пузыри, которые растут в сторону эктодермы в форме бокалов. Из стенки глазного бокала сформируются светочувствительные и нервные элементы сетчатки глаза, а из эктодермы в области глазных плакод и окружающей мезенхимы - все остальные структуры глаза. В силу этого зрительный нерв (II пара) не является типичным черепным нервом, а представляет собой специальное образование, связывающее сетчатку с головным мозгом, а сама сетчатка фактически является частью стенки нервной трубки, отделившейся от нее в процессе развития и выселившейся на периферию.
Одновременно с развитием конечного мозга начинают формироваться их ростральные выросты, из которых впоследствии образуются обонятельные луковицы и обонятельные тракты (I пара черепных нервов). Они также не являются типичными черепными нервами, а представляют собой проводниковый отдел обонятельного анализатора.
Конечный мозг с самого начала развития закладывается как парное образование в виде двух будущих полушарий. Внутри полушарий образуются парные полости - боковые желудочки мозга.
В период с 6-й до 12-й недели эмбриогенеза полушария головного мозга интенсивно растут, значительно опережая в росте все другие отделы головного мозга. В результате полушария, как плащ, покрывают эти другие отделы. Часть нейробластов мигрирует к поверхности стенки переднего мозгового пузыря, образуя в дальнейшем кору мозга.
К началу 4-го месяца внутриутробной жизни (т. е. чуть более, чем через 100 дней с момента начала развития организма) интенсивное деление нейробластов внутреннего плаща (коры большого мозга) приводит к тому, что у зародыша человека число нервных клеток уже приближается к их числу в коре у новорожденного.
Слой, в котором происходит размножение нейробластов, располагается наиболее глубоко, поэтому в процессе развития вновь образованные ней-робласты мигрируют наружу в мантийный слой, где они постепенно приобретают типичную для себя форму и занимают определенное положение в формирующейся коре мозга. Миграцию нейробластов направляют клетки радиальной глии, образующиеся из спонгиобластов эпендимы. Нейроны в процессе миграции как бы «скользят» вдоль отростков этих глиальных клеток из глубины мантийного слоя по направлению к поверхности мозгового пузыря. Нейробласты, мигрировавшие первыми, заполняют сначала
глубинные слои будущей коры, а клетки, образовавшиеся позднее, располагаются в более верхних слоях. Таким образом, кора головного мозга формируется как бы «изнутри кнаружи». При этом время образования нейрона определяет его принадлежность к определенным слоям коры, а значит и его
функции.
Созревание нейробластов происходит гораздо медленнее, чем их пролиферация (накопление количества), и продолжается после рождения в течение первых лет жизни. В коре мозга раньше всего созревают крупные пирамидные клетки ее нижнего этажа (проекционные нейроны), связывающие кору с нижележащими отделами головного и спинного мозга. Их можно наблюдать уже на 8-й неделе эмбриогенеза.
К 25-й неделе внутриутробного развития в корковой пластинке плаща (коры) можно различить цитоархитектонические слои. Рост и развитие плаща приводит к образованию борозд и извилин (складок) на поверхности конечного мозга, которые формируются на 28-30-й неделях. Окончательное завершение формирования рельефа полушарий наблюдается в постнатальном периоде примерно к 7-8 годам жизни ребенка. Последовательность и степень развития рельефа полушарий (борозд и извилин) программируется генетически, однако его строение в значительной степени зависит также от внешних влияний окружающей среды (соотношения благоприятных и неблагоприятных факторов существования, степени защищенности от влияния неблагоприятных условий, степени вынужденной активности индивидуума, информационной обогащенности среды и т. д.) и от состояния внутренней среды организма (уровня энергетического обмена, соотношения и уровня гормонов в крови и т. д.). Это обусловливает существенные индивидуальные различия в величине площади коры в разных долях и извилинах мозга.
Вначале поверхность больших полушарий гладкая. Первыми на 11 -12 неделе внутриутробного развития закладывается боковая борозда (Сильвиева), затем центральная (Ролландова) борозда. Довольно быстро происходит закладка борозд в пределах долей полушарий, за счет образования борозд и извилин увеличивается площадь коры.
Нейробласты путем миграции образуют скопления - ядра, формирующие серое вещество спинного мозга, а в стволе мозга - некоторые ядра черепно-мозговых нервов.
Сомы нейробластов имеют округлую форму. Развитие нейрона проявляется в появлении, росте и ветвлении отростков. На мембране нейрона образуется небольшое короткое выпячивание на месте будущего аксона - конус роста. Аксон вытягивается, и по нему доставляются питательные вещества к конусу роста. В начале развития у нейрона образуется большее число отростков по сравнению с конечным числом отростков зрелого нейрона. Часть отростков втягивается в сому нейрона, а оставшиеся растут в сторону других нейронов, с которыми они образуют синапсы.
В спинном мозге аксоны имеют небольшую длину и формируют межсегментарные связи. Более длинные проекционные волокна формируются позднее. Несколько позже аксона начинается рост дендритов. Все разветвления каждого дендрита образуются из одного ствола. Количество ветвей и длина дендритов не завершаются во внутриутробном периоде.
Увеличение массы мозга в пренатальный период происходит в основном за счет увеличения количества нейронов и количества глиальных клеток.
Развитие коры связано с образованием клеточных слоев (в коре мозжечка - три слоя, а в коре полушарий большого мозга - шесть слоев).
В формировании корковых слоев большую роль играют так называемые глиальные клетки. Эти клетки принимают радиальное положение и образуют два вертикально ориентированных длинных отростка. По отросткам этих радиальных глиальных клеток происходит миграция нейронов. Вначале образуются более поверхностные слои коры. Глиальные клетки принимают также участие в образовании миелиновой оболочки. Иногда одна глиальная клетка участвует в образовании миелиновых оболочек нескольких аксонов.
Таким образом, развитие головного мозга в пренатальный период происходит непрерывно и параллельно, однако характеризуется гетерохронией: скорость роста и развития филогенетически более древних образований больше, чем филогенетически более молодых образований.
Ведущую роль в росте и развитии нервной системы во внутриутробный период играют генетические факторы. Морфо-функциональное созревание нервной системы продолжается в постнатальный период.
Основное прибавление массы мозга приходится на первый год жизни ребенка. Увеличение массы мозга в постнатальный период происходит в основном за счет увеличения количества глиальных клеток. Количество нейронов не увеличивается, так как они теряют способность делиться уже в пренатальном периоде. Общая плотность нейронов (количество клеток в единице объема) уменьшается за счет роста сомы и отростков. У дендритов увеличивается количество ветвлений.
В постнатальном периоде продолжается также миелинизация нервных волокон как в центральной нервной системе, так и нервных волокон, входящих в состав периферических нервов (черепно-мозговых и спинномозговых).
Рост спинномозговых нервов связан с развитием опорно-двигательного аппарата и формированием нервно-мышечных синапсов, а рост черепно-мозговых нервов с созреванием органов чувств.
Таким образом, если в пренатальном периоде развитие нервной системы происходит под контролем генотипа и практически не зависит от влияния внешней окружающей среды, то в постнатальном периоде все большую роль играют внешние стимулы. Раздражение рецепторов вызывает афферентные потоки импульсов, которые стимулируют морфо-функциональное созревание мозга.
Под влиянием афферентных импульсов на дендритах корковых нейронов образуются шипики - выросты, представляющие собой особые постсинаптические мембраны. Чем больше шипиков, тем больше синапсов и тем большее участие принимает нейрон в обработке информации.
На протяжении всего постнатального онтогенеза вплоть до пубертатного периода, так же как и в пренатальный период, развитие мозга происходит гетерохронно. Так, окончательное созревание спинного мозга происходит раньше, чем головного мозга. Развитие стволовых и подкорковых структур раньше, чем корковых, рост и развитие возбудительных нейронов обгоняет рост и развитие тормозных нейронов. Это общие биологические закономерности роста и развития нервной системы.
Морфологическое созревание нервной системы коррелирует с особенностями ее функционирования на каждом этапе онтогенеза. Так, более раннее дифференцирование возбудительных нейронов по сравнению с тормозными нейронами обеспечивает преобладание мышечного тонуса сгибателей над тонусом разгибателей. Руки и ноги плода находятся в согнутом положении - это обусловливает позу, обеспечивающую минимальный объем, благодаря чему плод занимает меньшее место в матке.
Совершенствование координации движений, связанных с формированием нервных волокон, происходит на протяжении всего дошкольного и школьного периодов, что проявляется в. последовательном освоении позы сидения, стояния, ходьбы, письма и т.д.
Увеличение скорости движений обусловливается в основном процессами миелинизации периферических нервных волокон и увеличения скорости проведения возбуждения нервных импульсов. Более раннее созревание подкорковых структур по сравнению с корковыми, многие из которых входят в состав лимбической структуры, обусловливают особенности эмоционального развития детей (большая интенсивность эмоций, неумение их сдерживать связаны с незрелостью коры и ее слабым тормозным влиянием).
В пожилом и старческом возрасте происходят анатомические и гистологические изменения мозга. Часто происходит атрофия коры лобной и верхней теменной долей. Борозды становятся шире, желудочки мозга увеличиваются, объем белого вещества уменьшается. Происходит утолщение мозговых оболочек.
С возрастом нейроны уменьшаются в размерах, при этом количество ядер в клетках может увеличиться. В нейронах уменьшается также содержание РНК, необходимой для синтеза белков и ферментов. Это ухудшает трофические функции нейронов. Высказывается предположение, что такие нейроны быстрее утомляются.
В старческом возрасте нарушается также кровоснабжение мозга, стенки кровеносных сосудов утолщаются и на них откладываются холестериновые бляшки (атеросклероз). Это ухудшает деятельность нервной системы.
Органогенез головного мозга представлен на рис. . Антенатальное развитие ЦНС разделяется на три процесса: органогенез - формирование макроструктурных отделов мозга; гистогенез - развитие мозга от зарождения клеток; миграции дифференции. Конечным этапом является созревание ЦНС. Последнее характеризуется формированием синаптических контактов, нейронных сетей и миелинизацией, начинается на 36-й неделе внутриутробного развития и продолжается наиболее интенсивно до 5 лет. Созревание различных отделов ЦНС идет неравномерно. К концу антенатального периода лишь нервные клетки спинного мозга и глиальная ткань морфологически вполне развиты. Что касается полного структурного и функционального развития ЦНС, то оно завершается в постнатальном периоде. Для функциональной зрелости ЦНС большое значение имеет ми-елинизация проводящих путей, от степени развития которой зависят изолированное проведение в нервных волокнах и скорость проведения возбуждения. Именно миелинизация способствует увеличению объема головы с 37 см в окружности у новорожденного до 58-60 см - у взрослого.
Миелинизация различных путей в ЦНС обычно происходит в таком же порядке, в каком они развиваются в филогенезе. Например, миелинизация вестибулоспинального пути, являющегося наиболее примитивным, начинается с 4-го месяца, а руброспинального- с 5-8-го месяца внутриутробного развития. В спинном мозге и стволе мозга к моменту рождения основные проводящие пути миелинизированы. Исключение составляют пирамидный и оливоспинальный пути. В период внутриутробного развития начинается синтез большинства медиаторов, выделяемых в синапсах в ответ на раздражение клеток.
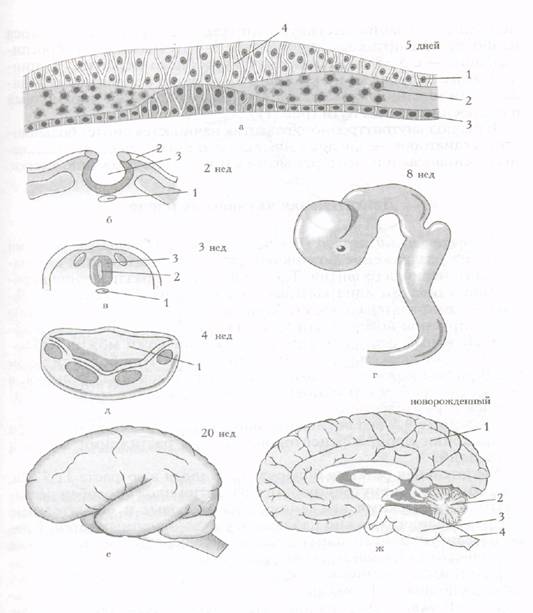
Рис. . Органогенез мозга:
а - нервная пластинка: 1 - эктодерма; 2 - мезодерма; 3 - энтодерма; 4 - нервная пластинка; б - нервный желобок: 1 - хорда; 2 - эктодерма; 3 - нервный желобок; в - нервная трубка: 1 - хорда; 2 - центральный канал; 3 - нервная трубка; г - образование мозговых пузырей; д - образование желудочков мозга: 1 - четвертый желудочек; е - формирование полушарий мозга; ж - увеличение массы и объема мозга: 1 - большие полушария; 2 - мозжечок; 3 - мост; 4 - продолговатый мозг
Наиболее интенсивное образование структур центральной нервной системы у плода происходит на 15-25-й день (табл. ). Структурное оформление отделов мозга тесно связано с происходящими в них процессами дифференцировки нервных элементов и установлением между ними морфологических и функциональных связей, а также с развитием периферических нервных аппаратов (рецепторов, афферентных и эфферентных путей и др.). К концу эмбрионального периода у плода обнаруживаются первые проявления нервной деятельности, которые выражаются в элементарных формах двигательной активности.
Функциональное созревание ЦНС происходит в этот период в каудо-краниальном направлении, т. е. от спинного мозга к коре головного мозга. В связи с этим функции организма плода регулируются преимущественно структурами спинного мозга.
К 7-10-й неделе внутриутробного периода функциональный контроль над более зрелым спинным мозгом начинает осуществлять продолговатый мозг. С 13-14-й недели появляются признаки контроля нижележащих отделов ЦНС со стороны среднего мозга.
Мозговые пузыри образуют полушария головного мозга. До четырехмесячного возраста плода их поверхность гладкая, затем появляются первичные борозды сенсорных полей коры, на шестом месяце - вторичные, а третичные продолжают формироваться и после рождения. В ответ на стимуляцию коры больших полушарий плода до 7 месяцев его развития никаких реакций не возникает. Следовательно, на этом этапе кора больших полушарий не определяет поведение плода.
На протяжении эмбрионального и фетального периодов онтогенеза строение и дифференцировка нейронов и глиальных постепенно усложняются.
Таблица
Развитие мозга в антенатальном периоде
| Зародыш/плод | Особенности развития мозга | |
| Возраст, нед. | Длина,мм | |
| 2,5 | 1,5 | Намечается нервная бороздка |
| 3,5 | 2,5 | Хорошо выраженная нервная бороздка быстро закрывается; нервный гребень имеет вид сплошной ленты |
| 4,0 | 5,0 | Нервная трубка замкнута; образовались 3 первичных мозговых пузыря; формируются нервы и ганглии; закончилось образование эпендимного, мантийного и краевого слоев |
| 5,0 | 8,0 | Формируются 5 мозговых пузырей; намечаются полушария большого мозга; нервы и ганглии выражены отчетливее (обособляется кора надпочечника) |
| 6,0 | 12,0 | Образуются 3 первичных изгиба нервной трубки; нервные сплетения сформированы; виден эпифиз (шишковидное тело); симпатические узлы образуют сегментарные скопления; намечаются мозговые оболочки |
| 7,0 | 17,0 | Полушария мозга достигают большого размера; хорошо выражены полосатое тело и зрительный бугор; воронка и карман Ратке смыкаются; появляются сосудистые сплетения (мозговое вещество надпочечника начинает проникать в кору) |
| 8,0 | 23,0 | В коре мозга появляются типичные нервные клетки; заметны обонятельные доли; отчетливо выражены твердая, мягкая и паутинная оболочки мозга; возникают хромаффинные тельца |
| 10,0 | 40,0 | Формируется дефинитивная внутренняя структура спинного мозга |
| 12,0 | 56,0 | Появляются общие структурные черты головного мозга; в спинном мозге видны шейное и поясничное утолщения; формируются конский хвост и концевая нить спинного мозга, начинается дифференцировка клеток нейроглии |
| 16,0 | 112,0 | Полушария покрывают большую часть мозгового ствола; становятся различимыми доли головного мозга; появляются бугорки четверохолмия; более выраженным становится мозжечок |
| 20-40 | 160-530 | Завершается формирование комиссур мозга (20 нед.); начинается миелинизация спинного мозга (20 нед.); появляются типичные слои коры головного мозга (25 нед.); быстро развиваются борозды и извилины головного мозга (28 - 30 нед.); происходит миелинизация головного мозга (36 - 40 нед.) |
Неокортекс уже у 7-8-месячного плода разделен на слои, но наибольшие темпы роста и дифференцировки клеточных элементов коры отмечаются в последние два месяца созревания плода и в первые месяцы после рождения ребенка. Пирамидная система, обеспечивающая произвольные движения, созревает позже, чем экстрапирамидная система, контролирующая непроизвольные движения. Показателем степени зрелости нервных структур является уровень миелинизации ее проводников ЦНС. Этот процесс в мозге эмбриона начинается на четвертом месяце внутриутробной жизни с передних корешков спинного мозга, подготавливая моторную активность плода; затем миелинизируются задние корешки, проводящие пути спинного мозга, афференты акустической и лабиринтной систем. В головном мозге основная часть проводниковых структур миелинизируется в первые 2 года жизни ребенка. Процесс миелинизации нервных волокон продолжается и в более старшем- даже зрелом возрасте.
Из рефлексов наиболее рано (7,5 недель) у плода появляется рефлекс на раздражение губ. Он хорошо выражен и локален. Рефлексогенная зона сосательного рефлекса к 24-й неделе внутриутробного развития значительно расширяется и вызывается со всей поверхности лица, кистей, предплечий. В постнатальном онтогенезе она вновь уменьшается до зоны поверхности губ.
На 8-й неделе раздражение периоральной области лица вызывает элементарную защитную реакцию - контралатеральное сгибание шеи, ведущее к отстранению стимулируемой поверхности головы от раздражителя (элементарный защитный рефлекс). При раздражении кожи можно наблюдать быстрое движение рук и туловища плода. При более сильном раздражении кожи часто возникает генерализованная реакция, в основе которой лежит одновременное сокращение мышц - сгибателей и разгибателей.
В возрасте 9,5 недель возможны двигательные реакции плода при раздражении проприорецепторов, т.е. при растяжении мышц и сухожилий.
Хватательная реакция обнаружена у плода в возрасте 11,5 недель. В поздние сроки внутриутробного развития плод способен реагировать мимическими движениями на вкусовые и обонятельные раздражители, что можно наблюдать у недоношенных детей. Кроме названных двигательных реакций наблюдается и спонтанная активность скелетной мускулатуры плода.
Рефлексы на тактильную стимуляцию кожи верхних конечностей появляются у плода к 11-й неделе. Наиболее четко кожный рефлекс в этот период вызывается с ладонной поверхности, проявляется он изолированными движениями пальцев. К этому же времени движения пальцев сопровождаются сгибанием запястья, предплечья, пронацией руки. К 15-й неделе стимуляция ладони приводит к сгибанию и фиксированию в этом положении пальцев, ранее генерализованная реакция исчезает. К 23-й неделе хватательный рефлекс усиливается, становится строго локальным. К 25-й неделе все сухожильные рефлексы рук становятся отчетливыми.
Рефлексы при стимулировании нижних конечностей появляются к 10-11-й неделе развития плода. Первым появляется флексорный рефлекс пальцев ног на раздражение подошвы. К 12- 13-й неделе флексорный рефлекс на это же раздражение сменяется веерообразным разведением пальцев. После 13 недель это же движение на раздражение подошвы сопровождается движениями стопы, голени, бедра. В более зрелом возрасте плода (22-23 недели) раздражение подошвы вызывает преимущественно флексию пальцев стопы.
К 18-й неделе появляется рефлекс сгибания туловища при раздражении нижней части живота. К 20-24-й неделе появляются рефлексы мышц брюшной стенки. К 23-й неделе у плода раздражением различных участков кожной поверхности можно вызвать дыхательные движения. К 25-й неделе плод может самостоятельно дышать, однако дыхательные движения, обеспечивающие выживание плода, устанавливаются только после 27 недель его развития.
Спонтанная активность мускулатуры плода характеризуется тремя основными формами:
1) тоническим сокращением мышц-сгибателей, обеспечивающим ортотоническую позу плода (согнутая шея, туловище и конечности), благодаря чему плод занимает в матке минимальный объем. Циркулирующие в полости амниона околоплодные воды и афферентная импульсация с проприорецепторов скелетных мышц поддерживают такое положение раздражением кожных рецепторов;
2) периодическими фазными (относительно короткими) сокращениями мышц-разгибателей, имеющими генерализованный характер. Шевеление плода начинается в 4,5 -5 месяцев беременности, и мать ощущает эти движения. Шевеления наблюдаются обычно 4-8 раз в течение часа. Их частота увеличивается при обеднении крови матери питательными веществами и при физических нагрузках беременной;
3) дыхательными движениями. Они начинаются на 14-й неделе внутриутробного развития и в конце беременности занимают 40-60 % всего времени. Частота дыхания очень высока: 40-70 в минуту. На 6-м месяце внутриутробного развития все основные части центральной регуляции дыхания уже достаточно зрелы, чтобы поддерживать ритмическое дыхание в течение 2-3 дней, а начиная с 6,5 -7-го месяца плод может дышать неопределенно долгое время.
Таким образом, рефлексы кожного, двигательного и вестибулярного анализаторов проявляются уже на ранних этапах внутриутробного развития. В поздние сроки внутриутробного развития плод способен реагировать мимическими движениями на вкусовые и обонятельные раздражения.
В течение трех последних месяцев внутриутробного развития у плода созревают рефлексы, необходимые для выживания новорожденного ребенка: начинает реализовываться корковая регуляция ориентировочных, защитных, пищевых и других рефлексов (у новорожденного эти рефлексы уже имеются); рефлексы с мышц и кожи становятся более локализованными и целенаправленными. У плода и новорожденного в связи с малым количеством тормозных медиаторов в ЦНС легко возникает генерализованное возбуждение - даже при очень небольших раздражениях. Сила тормозных процессов нарастает по мере созревания мозга.
Стадия генерализации ответных реакций и распространения возбуждения по структурам мозга сохраняется вплоть до рождения и некоторое время после него, но она не препятствует развитию сложных жизненно важных рефлексов. Например, к 21-24-й неделе хорошо развиты сосательный и хватательный рефлексы.
У плода уже на четвертом месяце хорошо развита проприоцептивная мышечная система, четко вызываются сухожильные и вестибулярные рефлексы, с этого же времени появляются лабиринтные и шейные тонические рефлексы положения: наклон и поворот головы сопровождается разгибанием конечностей той стороны, в которую повернута голова.
Рефлекторная функция у плода обеспечивается преимущественно механизмами спинного мозга и ствола головного мозга. Однако сенсомоторная кора уже реагирует возбуждением на раздражения рецепторов тройничного нерва на лице, рецепторов кожной поверхности конечностей; у 7-8-месячного плода в зрительной коре возникают реакции на световые стимулы, но в этот период кора, воспринимая сигналы, возбуждается локально и не передает значимые сигналы на другие (кроме двигательной коры) структуры мозга.
В последние недели внутриутробного развития у плода появляется цикличность сна: происходит чередование быстрого и медленного сна, причем быстрый сон занимает от 30 до 60% общего времени сна.
Дата добавления: 2018-02-28; просмотров: 1745; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!