Некоторые замечания по вопросам исторического развития динозавров 6 страница
Более поздние материковые отложения эры новой жизни (кайнозой) были изучены палеонтологами Академии наук СССР на юге страны, главным образом в Казахстане, и дали в руки ученых ископаемые остатки разнообразных представителей древних носорогов.
Между древними фаунами земноводных и пресмыкающихся и кайнозойскими фаунами млекопитающих прошел громадный отрезок геологического времени эры средней жизни (мезозой) с ее гигантскими ящерами — динозаврами. Материковые отложения этой эры на территории СССР не имеют широкого распространения, и для изучения их советские палеонтологи обратились к Центральной Азии. Вопреки существовавшему представлению об извечной пустынности Центральной Азии в мезозойскую эру советские палеонтологи, пользуясь тафономией, пришли к совершенно противоположному заключению о близости центральноаэиатской суши к морю, об ее низменном, заболоченном характере, обилии там воды и растительности. Эти заключения полностью подтверждены работами монгольской палеонтологической экспедиции Академии наук СССР, проведенной по приглашению правительства Монгольской Народной Республики в 1946—1949 годах. В Монголии были открыты богатейшие «кладбища» гигантских ящеров — динозавров — и древних млекопитающих. Полные скелеты этих животных привезены в Москву, препарируются и монтируются в Палеонтологическом музее. Директор музея профессор К. Флеров, являющийся и художником, занимается научно обоснованным восстановлением внешнего облика животных — созданием живописных и скульптурных реконструкций. По анатомическому строению скелета восстанавливается мышечная система, а затем и внешний облик животного.
|
|
|
Данные, полученные при тафономическом изучении местонахождений, дают К. Флерову возможность восстанавливать географические ландшафты, в которых обитали животные, и, таким образом, показывать этих животных в их жизненной обстановке, не отрывая представления о них самих от условий их существования.

Профессор И. Ефремов (слева), удостоенный в марте этого года Сталинской премии за научный труд «Тафономия и геологическая летопись», и директор Палеонтологического музея Академии наук СССР профессор К. Флеров. На нижнем снимке — скелет динозавра зауролофа, найденный советскими палеонтологами в пустыне Гоби

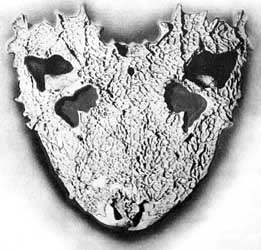
Уникальный череп лягушко-ящера лантанозуха, жившего около 200 миллионов лет наэад.

Скелет индрикотерия — гигантского носорога, жившего 30 миллионов лет назад. Этот скелет найден советскими палеонтологами в Казахстане. Внизу — внешний облик индрикотерия, восстановленный по строго обоснованным научным данным профессором К. Флеровым.
|
|
|

Источник:
Советский Союз. 1952. № 5. С. 34-35.
Вопросы изучения динозавров
(ПО МАТЕРИАЛАМ МОНГОЛЬСКОЙ ЭКСПЕДИЦИИ АКАДЕМИИ НАУК СССР)
Профессор И. А. Ефремов
Палеонтологическая экспедиция Академии наук СССР обнаружила на территории Монгольской Народной Республики крупнейшие местонахождения динозавров [cноска], различные по своему геологическому возрасту.
Эти местонахождения, подвергнутые тафономическому [cноска] изучению, доставили совершенно новые данные о захоронении динозавров и позволили подойти к выяснению причин формирования определенных жизненных группировок этих огромных ящеров.
С другой стороны, богатые результаты раскопок, выразившиеся в добыче полных скелетов, многих черепов и частей скелета как хищных, так и растительноядных дивоящеров, составили огромный материал, зачастую исключительно хорошей сохранности, открывающий новые возможности для изучения строения животных.
Таким образом, исследования Монгольской палеонтологической экспедиции важны тем, что в них впервые совмещено непосредственное изучение геологического строения местонахождений и заключенных в них остатков ископаемых животных.
|
|
|
В настоящей статье рассматриваются некоторые вопросы эволюции динозавров, наметившиеся при наблюдениях над новыми материалами и не освещавшиеся или неправильно истолковывавшиеся в прежних исследованиях.
Важнейшей чертой, характерной для дивоящеров и архозавров [cноска] вообще, является приспособление к двуногому типу передвижения. Все позднейшие четвероногие динозавры происходят от двуногих предков. Двуногий тип движения у пресмыкающихся появился очень давно — следы таких животных известны уже в каменноугольном периоде. Зарубежные исследователи обычно считали, что в развитии пресмыкающихся по труднообъяснимым («внутренним») причинам определились две главные линии развития: одна — двуногая, главенствующая у типичных рептилий, и другая — четвероногая, ведущая от самых примитивных форм через зверообразных пресмыкающихся к млекопитающим. И действительно у млекопитающих главенствует четвероногий тип передвижения, двуногие формы очень немногочисленны. Такое объяснение не может удовлетворить советского биолога, поэтому вопрос о появлении двуногого типа передвижения у пресмыкающихся, столь развитого в прошлые геологические эпохи, является одним из наиболее интересных в палеонтологии. Двуногость архозавров есть приспособление к определенным условиям существования, приспособление, проистекающее из особенностей строения организма пресмыкающегося.
|
|
|
Древние пресмыкающиеся с самого начала их возникновения обитали в громадных болотах и болотных лесах с густой растительностью, в прибрежных низменностях и дельтах. С эволюцией наземной растительности, все шире захватывавшей более высокие участки материков, первичные области обитания пресмыкающихся значительно расширились. Однако выход пресмыкающегося из затененных густой растительностью мест на открытые пространства, нагретые тропическим солнцем, был вовсе не так прост, как это нередко кажется исследователям, исходящим в своих представлениях из привычной человеку физиологии организма млекопитающего. Организм пресмыкающегося, не защищенный от перегревания ни специальной термоизоляцией, ни сложнейшим нервным механизмом регулирования постоянной температуры крови, на открытом пространстве днем подвергается опасности. Поэтому древнейшие четвероногие и зверообразные пресмыкающиеся, приспособляясь к обитанию на возвышенных, незатененных участках материка, дали в древнее, палеозойское время ряд ночных, или роющих форм, а еще большее число их стало обитать в реках с редкими выходами на сушу. У этих животных появились добавочные приспособления для регулирования температуры тела через теменной «глаз» при помощи желез внутренней секреции. Однако эти приспособления не разрешили главного биологического противоречия: при быстром передвижении организм сильно нагревается, и если он при этом не защищен еще от внешнего перегрева, то животное погибает. У четвероногих пресмыкающихся это противоречие было устранено только с появлением постоянной температуры крови и новых защитных покровов, иными словами, с возникновением млекопитающих. Млекопитающие обладают значительно более высокой энергией своего организма. Поэтому в своей приспособительной эволюции они могли обойтись без перехода к двуногому передвижению, т. е. типу движения с наименьшей затратой энергии, столь выгодному для менее активного организма пресмыкающегося. Архозавры обитали в местах, свойственных древнему периоду развития наземной жизни, — болотах и болотных низменных лесах. Естественно, что обширное распространение болотных низменностей с густой растительностью в мезозое определило развитие именно архозавров в их новых крупных формах — разнообразных динозавров.
Ведущим чувством пресмыкающегося является зрение, достигшее весьма высокого развития. Глаза пресмыкающихся и птиц совершеннее глаз млекопитающих, ведущими чувствами которых является обоняние и слух. Для ориентировки хищного пресмыкающегося, живущего в густой растительности, ему совершенно необходимо иметь высоко поднятую голову и в то же время возможно быстрее передвигаться. Для травоядного необходимы те же условия, чтобы избегать хищника. Оба эти основные требования условий существования разрешаются приспособлением к двуногому передвижению. Поэтому развитие множества двуногих пресмыкающихся — результат наилучшего приспособления к характерным и распространенным в мезозое обстановкам обитания. Фаунистический состав наиболее крупных местонахождений динозавров в Северной Америке, Европе и Восточной Африке теперь, после изучения захоронения динозавров в монгольских местонахождениях, приобретает новое освещение. Становится несомненным, что преобладающие формы в каждой фауне распределены зонально, соответственно условиям обитания.
Зауроподы — травоядные ящеротазовыс динозавры — были величайшими наземными животными всех геологических эпох. Для того чтобы объяснить их необыкновенные размеры, палеонтологи прибегали к самым фантастическим гипотезам. Распространен взгляд, что увеличенный гипофиз в мозгу зауропод свидетельствует о болезненном изменении этой важнейшей железы, вызвавшем гигантизм.
Водный образ жизни зауропод общепризнан. Однако существуют значительные расхождения в объяснении способа их питания. Маленькая голова и крайне слабое озубление этих животных не дают оснований допустить, что они вели хищный образ жизни, хотя существуют утверждения о питании зауропод мелкой рыбой, основывающиеся именно на характере озубления. Если попытаться объяснить строение зауропод, исходя из конкретных условий их существования, то мы придем к интересным сопоставлениям.
Возникновение зауропод в начале юрского периода и затем их расцвет вплоть до начала мелового периода явились ответом на особые, существовавшие в этот период условия внешней среды. Середина мезозоя — эпоха весьма низкого стояния материков, сходная с каменноугольным периодом, характерна развитием громадных прибрежных низменностей и лесных болот, с интенсивным углеобразованием. Именно эта часть мезозоя и совпадает с эпохой многочисленности зауропод, с их расцветом и всемирным распространением. Низкие берега материков, очень полого уходившие под уровень моря, образовывали громадную жизненную зону, несомненно обильную пищей. Однако эта зона долго не заселялась наземными животными. Именно для таких берегов характерны огромные приливные волны и очень широкая зона прилива — отлива. Таковы некоторые участки океанских берегов в настоящее время (на Зондских островах, на северо-востоке Австралии, во Флориде, Южной Америке, Африке и т. д.), где низменный берег постепенно переходит в морское дно. Ветры и волнения также создают здесь очень сильный накат, резко поднимающий уровень воды.
Более плоские материки среднего мезозоя в еще большей степени были подвержены приливным волнам. Исполинские волны ходили на большом пространстве, сразу поднимая уровень воды в прибрежных лагунах, озерках и болотах на несколько метров и силой своего напора сметая все, что не было приросшим ко дну. Поэтому древние четвероногие, обитатели прибрежий, могли жить только в затопленных лесах или на защищенном барьерными рифами берегу. Подобные две зоны захоронения мы видим в каменноугольную и в нижнепермскую эпохи для земноводных и пресмыкающихся: каменноугольные леса или «красные слои» Северной Америки (а также, вероятно, Приуралья), непосредственно связанные с рифами.
Эта прибрежная зона, распространившаяся на миллионы квадратных километров в эпоху развития низменностей, пустовала до появления зауропод. Гигантские животные в десятки тонн весом успешно сопротивлялись силе приливных волн, не тонули при подъемах воды и освоили новую, громадную зону обитания, где им не было конкурентов. В свете высказанных соображений становится понятным развитие у зауропод больших когтей, необходимых для цепляния за грунт. Серии следов зауропод в Северной Америке найдены в вполне подходящих отложениях мелкой лагуны, с мягким, но не топким дном. Приспособительная эволюция зауропод во времени показывает нам картину постепенного освоения все больших глубин прибрежья, возможно, отчасти связанную с постепенным повышением материков и более резким профилем берегов.
Так, если юрские зауроподы (бронтозавры) могли обитать на глубине до трех метров, а верхнеюрские (диплодок) — до четырех-пяти метров, то нижнемеловые брахиозавры могли кормиться на глубине до восьми метров. Развитие зауропод наглядно показывает, как прежний двуногий тип строения скелета (платеозавр) вновь становится четвероногим, но все еще с ведущим значением задних конечностей, более длинных и более тяжелых (диплодок), и, наконец, превращается в жирафоподобный, со значительно сильнее развитыми передними конечностями, покатой назад спиной и сильно поднятой шеей (брахиозавр).

Зауроподы в приливной волне (первая зона)
Реконструкция К. К. Флерова
Гигантизм зауропод не был случайным, незакономерным или болезненным явлением, а явился приспособлением к такой обстановке обитания, в которой могли жить только гиганты. Увеличение роста и, вероятно, также ускорение его для того, чтобы молодые животные быстрее достигали размеров, минимально необходимых для жизни в особых условиях существования, конечно, требовали усиленного действия гипофиза. Большой гипофиз был нужен для достижения громадного роста, именно этим и объясняется увеличение гипофиза у зауропод, а вовсе не тем, что увеличение его (от неизвестных причин) вызвало увеличение роста.
Вопрос о роде питания зауропод не может быть пока разрешен из-за недостаточных сведений о флоре того времени. Однако можно заранее сказать: челюстной аппарат зауропод так непропорционально мал и слаб по сравнению с размерами этих животных, что растительная пища должна была быть чрезвычайно мягкой и питательной. Вероятно, в мелких лагунах и болотах середины мезозоя существовала еще неизвестная нам растительность, может быть, типа питательных студенистых водорослей. Питание рыбой представляется мне невероятным, так как оно требует и более сильного озубления и значительно большей подвижности, чем это показывает скелет зауропод, с его малоразработанными сочленениями. Предположение, что зауроподы занимались ловлей мелкой рыбешки при помощи громадной шеи с ее полуметровыми позвонками, равносильно допущению стрельбы из пушки по воробьям. Такая нелепость не могла существовать в природе. Возможно, что какие-нибудь планктонные животные, икра и личинки могли образовывать в лагунах значительные скопления и служить едой зауропод. Принимая во внимание обилие прибрежных беспозвоночных в мезозое, предположение об этом способе питания зауропод, как своего рода четвероногих китов, не заключает в себе ничего невероятного.
Сокращение затопляемых низменных прибрежий с середины мелового периода вызвало сужение областей обитания зауропод. Для каждого рода крупных наземных позвоночных площадь обитания должна быть очень большой. Сокращение ее размеров не приводит к простому уменьшению численности животных, но влечет за собой вымирание.
И если в середине мезозоя на материковых побережьях лишь сравнительно небольшие участки были недоступны постоянным затоплениям и приливным волнам, то в кайнозое и особенно в настоящее время стали редкими, наоборот, пониженные участки. Современные ландшафты ничего уже не говорят нам о некогда обширном распространении низменных побережий, вызвавших к жизни чудовищных зауропод.
Места обитания зауропод располагаются на самом внешнем крае материка и несколько выдвигаются в море. Более высокие области тех же низменных побережий, удаленные от непосредственного соприкосновения с морем, обладают зональным распространением различных динозавров.
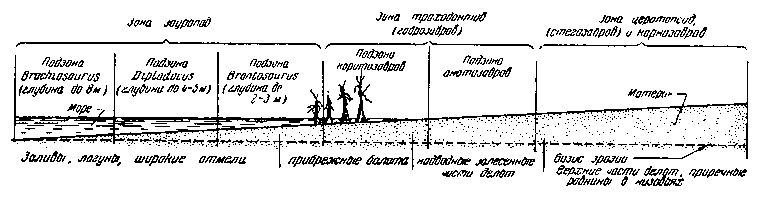
Схема основных зон обитания динозавров в мезозое. Деревья обозначают границу открытого побережья и низменных прибрежных лесов
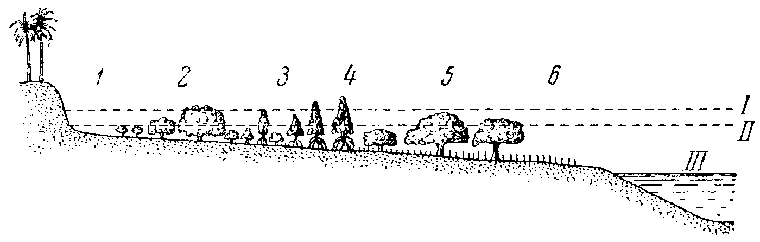
Схематический профиль затопляемого морского побережья у Танга (Восточная Африка). Тип закрытого растительностью побережья (зона II). I —высший уровень прилива, II — нормальный прилив, III — низший уровень отлива. Пояса мангровой растительности: 1 — пояс, лишенный растительности; 2 — прибойная зона высшего уровня (пояс с авиценниями); 3 и 4 — высококорневые мангровики (пояс цериопсов и ризофор); 5 — прибойная зона нормального уровня (массивные авиценнии и соннерации); 6 — границы древесной растительности (открытая низменность)
В низовьях дельт, защищенных от приливов болотными лесами, в бесчисленных протоках окружающих болот обитали различные виды утконосых динозавров (траходонтов или гадрозавров), занимавших следующую за зауроподами зону. Зона траходонтов распадается на две подзоны. Одну, с более глубокими водами, занимали коритозавры, паразауролофы и вообще все формы с дополнительными воздушными камерами в носовых полостях и сложным устройством дыхательных путей, с узкими рылами. Вторая, мелководная подзона, служила местом обитания широкорылых анатозавров, в самых общих чертах своей биологии похожих на современных бегемотов.
Характерно, что обитатели обеих рассмотренных зон — зауроподы и траходонты — не обладали никакими приспособлениями как для активной, так и для пассивной зашиты. Из этого следует, что обитание в обеих зонах само по себе предохраняло травоядных от непосредственного соприкосновения с хищниками.
Следующая, третья зона, заходящая далеко в материк, захватывает внутренне-материковые окраины прибрежных болотистых лесов и речных низовий и еще не поддается дальнейшему разделению. В этой зоне господствуют хищники — громадные карнозавры, и все обитающие здесь травоядные несут тяжелое панцырное вооружение, появление которого не случайно. В юре и нижнем мелу здесь жили стегозавры с громадными гребнями вдоль спины. Позднее они заменяются различными анкилозаврами, покрытыми тяжелыми костными плитами (сколозавры и анкилозавры). Здесь же обитали рогатые быкоподобные динозавры — цератопсиды.
Карнозавр в бою с цератопсами (третья зона)
Реконструкция К. К. Флерова
Распространенные представления о постоянных нападениях карнозавров на зауропод несомненно ошибочны. Не говоря уже о том, что зона обитания зауропод была мало доступна для карнозавров, прямое нападение на зауропода, даже вышедшего, скажем, для кладки яиц, в «чужую» зону, могло оказаться успешным лишь в очень редких, исключительных случаях. Траходонты, вероятно, иногда служили пищей карнозаврам, но и тут нападение на чрезвычайно массивного и сильного траходонта могло окончиться гибелью хищника. Не случайно наиболее крупные карнозавры (тиранозавры) появились не в период наибольшего расцвета зауропод, а значительно позже.
Рассмотренная выше зональность обитания динозавров отражена в большинстве местонахождений. Именно так группируются остатки преобладающих видов животных.
Громадные скопления остатков зауропод в верхней юре и нижнем мелу Уайоминга в Северной Америке или Тендагуру в Восточной Африке отражают захоронение первой, зауроподовой зоны. Остатки хищников, или стегозавров, там очень редки. Верхнемеловые местонахождения США показывают или скопления костей утконосых динозавров, или совместное нахождение остатков рогатых панцырных и хищных дивоящеров. В Европе и Америке юрские хищники найдены вместе со стегозаврами.
Местонахождения Монголии своеобразны: на фоне очень редких остатков зауропод в Нэмэгэту часто встречаются хищные карнозавры и утконосые траходонты. Поэтому местонахождение Нэмэгэту нужно рассматривать как пограничное между зоной траходонтов и зоной карнозавров, со случайно попадавшими сюда зауроподами.
Дата добавления: 2020-04-25; просмотров: 195; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!