Некоторые замечания по вопросам исторического развития динозавров 2 страница
Другим, очень важным вопросом, игнорировавшимся в прежних исследованиях, становится вопрос о размножении динозавров.
Исследования захоронения пресловутых яиц динозавров в главном и по существу единственном их местонахождении — Баин-Дзак (Шабарак-Усу, Джадохта), проведенные нашей экспедицией, показали, что эти яйца скорее принадлежат не динозаврам, а нескольким видам крупных черепах. Нужно думать, что подлинных яиц динозавров еще неизвестно в геологической летописи.
Однако при колоссальной численности динозавров, отраженной в громадных местонахождениях, сохранивших лишь ничтожную долю истинного количества животных, в прошлом должно было существовать исполинское количество кладок яиц. Если три вида речных черепах в реках Южной Америки откладывают ежегодно сотни тысяч и миллионы яиц, буквально начиняя ими речные отмели, если яйца крокодилов в мостах изобилия этих животных встречаются на каждом шагу на речных островах, то, безусловно, кладки яиц динозавров исчислялись миллиардами. Совершенно невероятно, чтобы на всем земном шаре, за шестьдесят миллионов лет господства и расцвета динозавров, из этих миллиардов яиц не получилось их частого захоронения и даже обилия в некоторых местонахождениях. Повидимому, яйца динозавров не имели твердой скорлупы и потому отсутствуют в геологической летописи. Также весьма возможно, что часть динозавров рождала живых детенышей, особенно те, зоны обитания которых неблагоприятны для сохранения яиц, как, например, у зауропод, для которых необходимы специальные убежища размножения, а откладывание яиц могло происходить лишь в чужой, более высокой зоне.
Если динозавры обладали яйцами в мягкой скорлупе, требующими большой влажности для сохранения, то размножение этих пресмыкающихся могло происходить только в глубине болот. Поэтому возможны были сезонные миграции динозавров в места размножения и связанная с этим частая гибель в чужих биотопах (как, например, зауропод).
Твердая скорлупа яиц, при постоянном обитании динозавров в 6олотистых низменностях, была им по существу ненужной.
Вместе с тем громадное количество яиц, ежегодно откладывавшихся пресмыкающимися в различные времена года, составляло десятки тысяч тонн питательной биомассы и не могло остаться неиспользованным в приспособительной эволюции наземных позвоночных. Вполне вероятно, что на пище из яиц размножались и благоденствовали различные примитивные млекопитающие. С этой точки зрения было бы очень интересно попытаться найти следы яйцеядных адаптаций среди палеоценовых млекопитающих, во многом сохранивших приспособительные структуры их меловых предков (рис. 3).
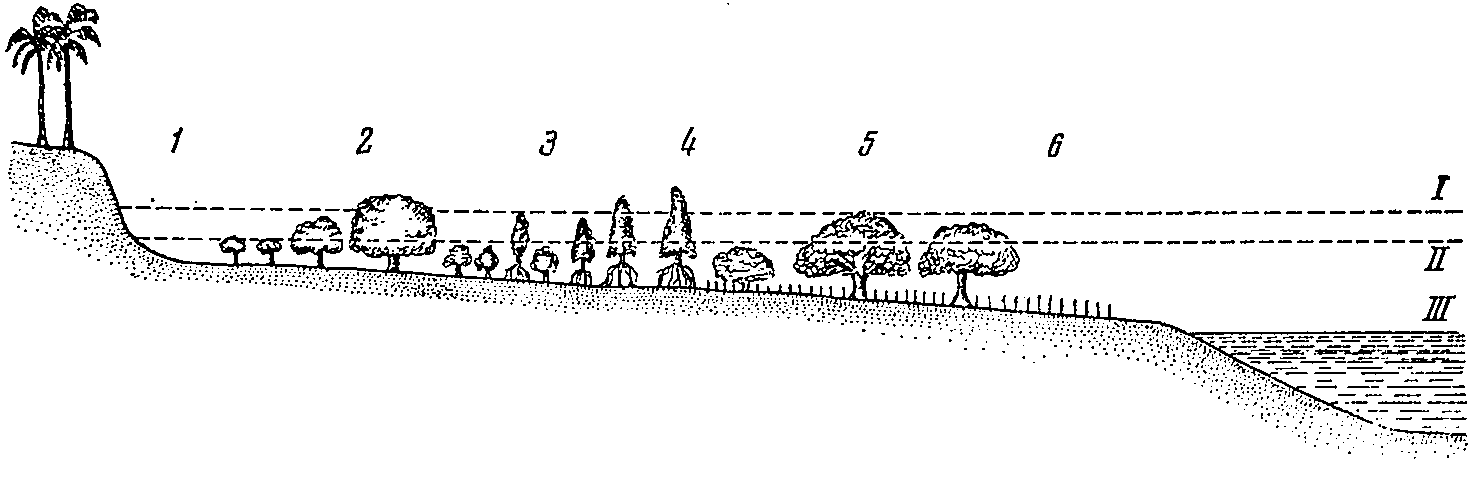
Рис. 3. Схематический профиль затопляемого морского побережья у Танга (Восточная Африка) (из Алехина, 1950). Характерна смена поясов мангровой растительности. Тип закрытого растительностью побережья (зона II).
1 — пояс, лишенный растительности; 2 — пояс с господством Avicennia; 3 — пояс Ceriops; 4 — пояс Rhizophora; 5 — пояс Sonneralia; 6 — граница распространения прибрежной растительности (открытая низменность). I — высший уровень прилива; II — нормальный прилив; III — низший уровень отлива.
Откладывание яиц в сухих местностях, где требовалась твердая скорлупа, вместе с тем сделало бы кладки более доступными для мелких млекопитающих и обусловило бы частое поедание кладок. Этого противоречия не существовало в пермскую эпоху, когда некоторые рептилии могли приспособиться к откладыванию твердых яиц в безопасных от затопления местах. Область,. находящаяся среди огромных болот, где динозавры откладывали свои мягкие яйца, была малодоступна млекопитающим, но зато подвержена затоплениям и связанной с ними частой гибели кладок. Это условие должно учитываться при изучении размножения динозавров.
Гораздо более серьезное противоречие — это назревшая необходимость заботы о потомстве. Крупные динозавры обладали уже настолько усложненной организацией, что их развитие не требовало длительного эмбрионального или постэмбрионального периода, и только что вылупившиеся молодые животные должны были быть довольно беспомощными.
Если представить себе крохотного, только что вылупившегося из яйца детеныша крупного карнозавра — двуногое существо с огромной тяжелой головой, почти без передних конечностей, то станет совершенно ясна его беспомощность и неспособность к самостоятельному существованию.
Известно, что двуногая походка требует развития сложных рефлексных систем для автоматизации равновесия, поэтому такая походка требует обучения на первых шагах вступления в жизнь. Обучение двуногой ходьбе было особенно важно для карнозавров, так как другие двуногие динозавры, как, например, траходонты, могли помогать себе передними конечностями, довольно сильными и снабженными плавательными перепонками. Карнозавры же, помимо двух ног, третьей и последней опорой имели хвост и представляли собою своеобразные треножники.
Не нужно забывать, что основным жизненным приспособлением зауропод является их гигантская величина, следовательно, маленькие зауроподы нежизнеспособны и требуют также охраны со стороны родителей.
Поэтому очевидно, что нормальное развитие динозавров может быть обеспечено при одном из трех следующих условий: а) громадных, хорошо защищенных яйцах, с удлиненным периодом развития (черепахи), б) очень большом числе мелких яиц, с массовой гибелью детенышей (крокодилы), в) заботе о потомстве, включая кормление и обучение, а не только охрану.
Громадные яйца несовместимы с прогрессирующим сужением таза при двуногом передвижении. Огромная смертность и мелкие детеныши очень невыгодны для крупных животных, не обеспечивают развития усложненного организма и замедляют вступление животного в нормальный биоценоз. Реальным разрешением вопроса является только забота о потомстве, необходимость которой вступает в противоречие со слабым развитием мозга у всех без исключения динозавров.
Это противоречие так и осталось неснятым и явилось самым слабым местом во всей эволюции динозавров. Только развитие мозга у млекопитающих обеспечило дальнейшее усложнение организма позвоночных путем сильного удлинения постэмбрионального развития, которое стало возможным при долгой, сложной и тщательной заботе о потомстве. Микроцефалы, подобные динозавру, ни при каких условия не могли бы проявить подобной заботы о потомстве. У птиц забота сводиться в основном к кормлению и очень сильно облегчается периодом неподвижности детенышей, чего не бывает у четвероногих.
Противоречие между развитием мозга и необходимостью заботы о потомстве стало одним из основных, остановивших дальнейшую эволюцию динозавров и приведших к вымиранию.
Приведенные соображения показывают, что понимание путей эволюции, расселения и причин вымирания динозавров может быть достигнуто только в результате многостороннего анализа строения организма динозавров и его взаимоотношения со средой обитания.
Исследования частных приспособительных структур в скелете динозавров находятся в зачаточном состоянии. Еще очень мало черт строения динозавров получило свое функциональное объяснение.
Между тем собранные Монгольской экспедицией костные материалы показывают черты анатомического строения, соответствующие функции в тех определенных условиях существования, которые наметились при исследовании местонахождений. Изменение условий существования, отмеченное сменой фаций и фаун в открытых экспедицией местонахождениях, неуклонно влечет за собой изменение приспособительных структур и возникновение новых противоречий между организмом, с его анатомо-физиологическими возможностями, и средой обитания.
Древнее появление двуногости у архозавров обусловило, вне всякого сомнения, развитие ряда специфических анатомических структур. Изучение их на монгольских находках, отличающихся полнотой и хорошей сохранностью, позволит проследить разнокачественные приспособления двуногого пресмыкающегося к разным условиям существования. Динозавры, как представители далеко прогрессировавших двуногих архозавров, особенно монгольские верхнемеловые формы, должны обладать резким выражением приспособительных структур, поддающимся функционально-морфологическому анализу.
Так, например, траходонтные травоядные динозавры отчетливо выказывают прогрессирующее приспособление к питанию сравнительно жесткой растительностью. Это прежде всего развитие озубления из многих рядов зубов, давно известное для траходонтов. Однако до сих пор неизвестно, что эти гипсодонтные зубы при взаимодействии дают длинную рабочую поверхность, с острым и зазубренным режущим внутренним краем, составленным из внутренних эмалевых стенок зубов верхне-внутреннего ряда. Эмалевые стенки других зубов образуют на рабочей поверхности сетку — ряд ячей из эмалевых перегородок, функционально аналогичную сложным гипсодонтным зубам млекопитающих.
Такое строение зубов траходонтов противоречит, на первый взгляд, их плавательным перепонкам на передних конечностях и водному образу жизни. Возможное объяснение этого противоречия тем, что при питании болотной растительностью на зубы траходонтов попадало большое количество земли, способствовавшей быстрому истиранию зубов, не соответствует строению их морды, отнюдь не приспособленной для глубокого захвата растений вместе с корнями, как это делают бегемоты и свиньи. Вероятно, что траходонты представляли собой дальнейшее развитие игуанодонтового экологического типа. Игуанодонты, двуногие травоядные нижнего мела, эволюционировали в сторону обитания в третей, более сухой зоне. Однако развитие гигантских карнозавров, вероятно, вынудило примитивных траходонтов отступить снова во вторую зону, в связи с чем у них выработался ряд мелких адаптивных приспособлений к водному обитанию, как, например, плавательные перепонки на передних конечностях. Однако сложные зубы впоследствии, при сокращении второй зоны к концу мела и появлении в ней более жесткой растительности, обеспечили анатозаврам процветание.
В соответствии с новым способом питания развивается мощный короноидный отросток на нижней челюсти и тяга приводящих мышц осуществляется за счет группы мышц массетер-темпоралис. Птеригоидные мышцы редуцируются, и вместе с ними редуцируются птеригоидные нисходящие отростки, что дает свободу нижней челюсти. Длинные квадратные кости с низко опущенными челюстными мыщелками допускают широкое раскрытие рта и позволяют нижней челюсти совершать передне-задние жевательные движения. Зубы находятся только в заднем конце челюстей, что повышает эффективность их работы. У анатозавров развиваются целые батареи зубов, до пятисот на каждой стороне челюсти. Соответственно лучшему приспособлению к новым условиям существования, анатозавры очень многочисленны в верхнем мелу и существуют до конца эпохи.
Развитие сложной зубной системы у траходонтных динозавров привело к увеличению размеров черепа, который становится более высоким и несет много мощных мышц. Для удержания тяжелой головы при двуногом передвижении у траходонтов появляются необычайно мощные мышечные крепления длинных и широких мышц спины. Остистые отростки крестцовых позвонков достигают полуметра высоты, указывая на соответственную толщину прикреплявшихся здесь мышц. Поэтому разгибатели спины у траходонтов должны были обладать совершенно исключительной силой. Повидимому, при движении позвоночник траходонтов подвергался сильному прогибанию вниз в середине позвоночного столба, с резким подъемом головы и хвоста, как это совершается у современных кенгуру. Невральные дуги позвонков — сильно развитые и высокие, с большими, приподнятыми вверх диапофизами, которые, вместе с остистым отростком образуют в каждом позвонке три радиально расходящихся и направленных вверх массивных отростка. Совершенно такое же строение мы встречаем у современных двуногих млекопитающих — тушканчиков, у которых оно служит укреплением позвоночника при несении свободно висящей передней части тела со сравнительно хорошо развитыми передними конечностями. У карнозавров с их крошечными передними конечностями передние отростки развиты слабее. Встреченные в наших материалах окостенения сухожилий также говорит о постоянной нагрузке позвоночника траходонтов на прогиб. Все это свидетельствует о полусогнутой, а не вертикальной походке птицетазовых двуногих динозавров. Повидимому, более выпрямленным положением тела при движении отличались крупные карнозавры, которые вообще представляют крайнее развитие наиболее подвижного двуногого пресмыкающегося.
Особенностью, указывающей на различие в характере движения задних конечностей у карнозавров и траходонтов, служит положение четвертого трохантера на бедренной кости. У всех без исключения траходонтов (так же как и игуанодонтов) четвертый трохантер развит в виде мощного костного гребня в несколько сантиметров высоты и располагается почти точно посредине длины бедра. Здесь, несомненно, прикреплялись чрезвычайно мощные пучки хвостобедренных (каудофеморальных) мышц.
Комбинация мощных мускулов и низкого приложения их тяги давала очень большую силу отведения конечности назад и отталкивания от почвы. Вместе с тем эти же мышцы служили для поддержки тулоговища при наклоне головы вперед и вниз, когда создавался перенос центра тяжести вперед. Огромная толкающая сила задних конечностей животного, полученная в результате этого приспособления, находилась в противоречии с подвижностью ног. Размах бедра, вынос задней конечности вперед при шаге получились весьма ограниченными. Вообще чрезвычайно грубый скелет траходонтов, с плохо развитыми сочленениями, столь контрастирующими с как бы шлифованными сочленениями карнозавров, с перегруженной, слишком тяжелой передней половиной туловища, с непропорционально небольшим тазом и тяжелыми задними ногами, по массивности не уступающими ногам зауроподов, показывает, что внешний облик животного был совершенно своеобразен и мало пригоден для сухопутного передвижения. Вероятно, животное выходило из воды лишь в очень редких случаях, возможно, только для кладки яиц, и вне воды было неуклюжим и медленно двигающимся.
Совершенно аналогичное развитие четвертого трохантера наблюдем у зауропод, с той лишь разницей, что прикрепление хвостобедренных мышц приходилось на несколько бoльшую площадь и гребень трохантера не был столь высоким. Следует считать, что характер движения задних конечностей у зауропод и траходонтов был очень сходен. Обязано ли это сходство общности приспособления на начальных стадиях развития или общности происхождения — подлежит выяснению.
У карнозавров четвертый трохантер развит значительно слабее и расположен гораздо выше — на уровне первой трети длины бедра. Другие трохантеры вынесены высоко вверх и образуют с наружной стороны шейки бедра мощный вырост, поднимающийся выше головки кости. Все главные двигающие бедром мускулы прикреплялись на верхней трети удлиненной кости, что при проигрыше в силе обеспечивало большой размах ноги, а следовательно, и быстроту движения для пресмыкающихся, у которых мышцы не дают очень частых повторных сокращений. Большой размах обеспечивался и поднятием трохантерного бугра выше оси вращения головки, так что получалось как бы второе плечо рычага. Однако получившееся ослабление тяги хвостобедренных мускулов неизбежно должно было привести к редукции передних конечностей для предельного облегчения передней части туловища, при двуногой походке находящейся на весу. Это же ослабление хвостобедренных мышц показывает, что карнозавры не могли быть столь же быстрыми пловцами, как траходонты. На монгольских находках отмечается чрезвычайное сужение таза карнозавров в полном соответствии тому, что максимальное сближение головок бедренных костей наиболее выгодно при вертикальном двуногом передвижении.
Очень интересно развитие больших валикообразных суставных площадок на шейках бедренных костей. Помимо головки, расширенной и слегка уплощенной, входящей глубоко в ацетабулярную яму и ориентированной в вертикальной плоскости, бедро карнозавра снабжено длинной и широкой суставной площадкой, занимающей всю верхнюю оконечность кости от головки до трохантерного внешнего угла. Эта площадка располагается на утолщенной верхушке кости и ориентирована в горизонтальной плоскости. Трение в этом суставе происходило между описанной площадкой и верхней половиной стенки ацетабулярной ямы. Основной трохантер выдвинут вперед. Аналогичное, но гораздо слабее выраженное устройство наблюдается у современных тушканчиков, у которых с внутренней стороны головки образуется суставная поверхность, заходящая за шейку бедра. Это рассматривается советскими зоологами (Виноградов, 1937) как приспособление к постоянному передвижению прыжками с одновременным участием обеих ног. Такой способ движения, причем еще сильнее специализированный (судя по гораздо большему развитию дополнительных суставных площадок), можно допустить для мелких и легких карнозавров типа горгозавров. Однако точно такое же строение мы видим у крупных монгольских карнозавров. Допустить передвижение исключительно прыжками животного, весящего несколько тонн (около 7 т у крупных) очень трудно. Устройство конечностей карнозавров функционально еще не ясно.
Увеличение головы, являющейся единственным орудием нападения, не компенсируется, как у траходонтов, развитием гигантских мышц спины. Судя по строению таза и позвонков, разгибательные мышцы спины у крупных карнозавров слабее, чем у траходонтов. Зато таз, особенно у крупных карнозавров, развит сильнее, чем у траходонтов, превосходит размеры черепа и представляет мощную конструкцию — основной несущий центр треножника, на котором сидит все тело. Замечательно развитие спинного гребня из сходящихся вверху подвздошных костей (сильно удлиненных), глубоких ацетабулярных ям и массивного молоткообразного утолщения на срощенном симфизе лобковых костей.
Шейные позвонки карнозавров уплощены и расширены, обладают косыми суставными площадками центров, обеспечивающими при поднятой голове прочную фиксацию позвоночника. Вызывавшаяся этим малая подвижность головы должна была обусловливать совершенно особый способ нападения карнозавров. Повидимому, эти хищники производили нападение сверху, ударяя всем весом туловища, поворачивавшегося в тазобедренных сочленениях (рис. 4). Возможно, что развитие дополнительных суставов на бедренных костях связано с этим способом нападения. Во всяком случае интересно, что все панцырные травоядные динозавры защищены сверху.
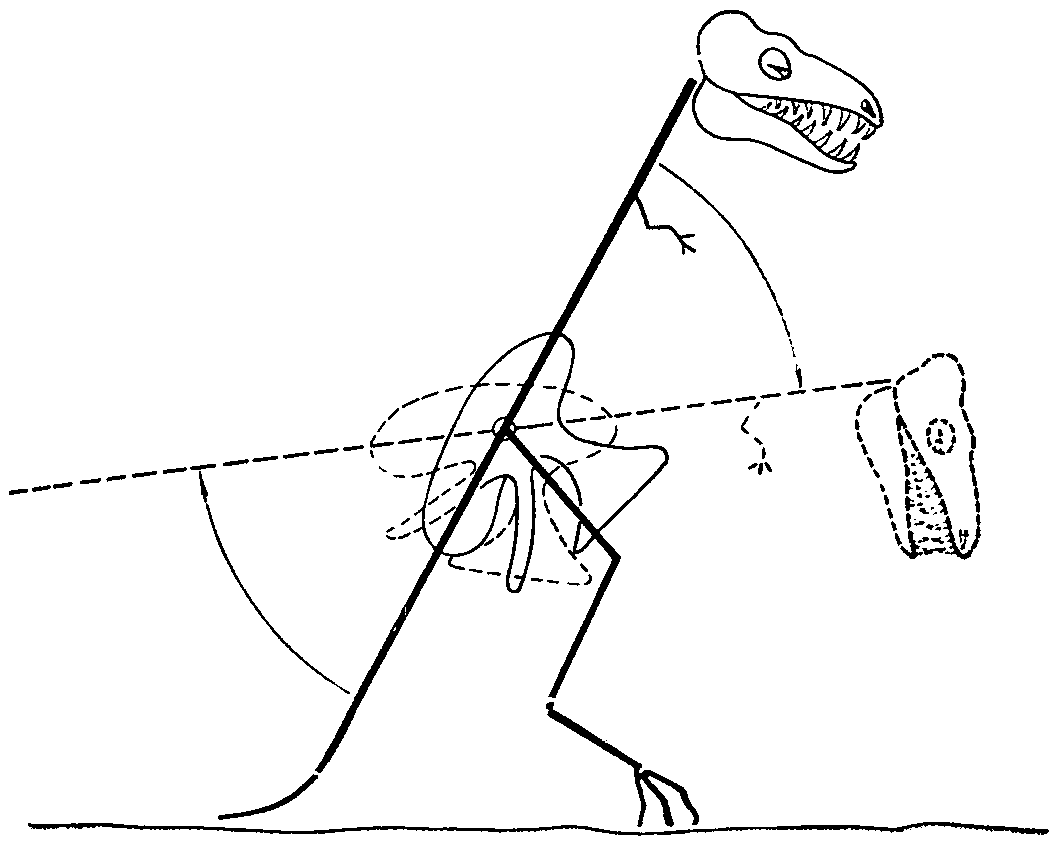
Рис. 4. Схема способа нападения крупного карнозавра. (Примерно 1/80 натуральной величины животного. Полная длина хвоста для уменьшения рисунка не показана).
Как разрешаются противоречия в сужении таза и произведении потомства, малой подвижности головы и активном нападении? Ответить на эти интересные вопросы должны последующие детальные исследования.
Следует отметить, что орбиты карнозавров располагаются в выступающих наружу углах черепа. Подобное устройство орбит позволяет глазам повертываться прямо вперед и, следовательно, обладать стереоскопическим зрением, очень важным для хищника при расчете прыжка или удара. Сходным бинокулярным зрением обладают крупные хищные млекопитающие.
В соответствии с развитием гигантской головы у поздних хищных карнозавров произошла редукция передних конечностей — как необходимая корреляция для уравновешивания передней части туловища. Передние конечности, подобные таковым у траходонтов, обусловили 6ы значительную перегрузку переднего конца тела, которая должна была сделать невозможным двуногое передвижение.
Так как передние конечности карнозавров не могли помогать при еде, в черепе больших карнозавров появляется кинетичность — внутренние сочленения, позволявшие передней части черепа двигаться относительно задней. Повидимому, это приспособление помогало проталкиванию в глотку больших кусков мяса без участия передних конечностей, обязательного у всех других хищников.
Дата добавления: 2020-04-25; просмотров: 210; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!