Некоторые замечания по вопросам исторического развития динозавров 1 страница
Монгольская палеонтологическая экспедиция обнаружила на территории МНР многочисленные остатки фауны динозавров в различных по геологическому возрасту местонахождениях. Эти местонахождения, подвергнутые тафономическому изучению, доставили совершенно новые данные о захоронении динозавров и позволили подойти к выяснению причин формирования определенных палеофаунистических группировок.
Кроме того, богатые результаты раскопок, выразившиеся в добыче полных скелетов, многих черепов и частей скелета как хищных, так и растительноядных динозавров, составили огромный материал, зачастую исключительно хорошей сохранности, открывающий новые возможности для изучения строения этих животных.
Даже при первом знакомстве с новыми материалами бросается в глаза много особенностей, игнорировавшихся или упущенных в основных работах (Huene, 1917; Gilmore, 1930, 1936; Nopcsa, 1930; Mook, 1917; Osborn, 1921 и др.) по ископаемым позвоночным Северной Америки и Европы.
Исследования Монгольской палеонтологической экспедиции АН СССР [ценны тем], что в них впервые в отношении мезозойских наземных позвоночных непосредственное изучение геологического [образова]ния местонахождений совмещено с морфолого-палеонтологическим анализом заключенной в них фауны.
В настоящей статье рассматриваются некоторые вопросы эволюции динозавров, наметившиеся при непосредственных наблюдениях над тафономией и костными материалами и не освещавшиеся или неправильно истолковывавшиеся в прежних исследованиях.
Как известно, важнейшей чертой, характерной для динозавров и архозавров вообще, является приспособление к двуногому типу передвижения. Все позднейшие четвероногие динозавры происходят от двуногих предков. Двуногий тип движения у пресмыкающихся появился очень давно — следы двуногих форм известны уже в каменноугольном периоде. Зарубежными исследователями обычно принималось, что в развитии пресмыкающихся, по причинам внутреннего порядка, определились две главные линии развития — одна двуногих животных, главенствующая у типичных рептилий и другая — четвероногих животных, ведущая от самых примитивных форм через зверообразных пресмыкающихся к млекопитающим. И действительно, у млекопитающих главенствует четвероногий тип передвижения, двуногие формы очень немногочисленны. Такое «объяснение» не может удовлетворить советского биолога, поэтому вопрос о появлении двуногости у пресмыкающихся, столь развитой в прошлые геологические эпохи, является одним из наиболее интересных в палеонтологии.
Двуногость архозавров есть приспособление к совершенно определенным условиям существования, приспособление, проистекающее из особенностей строения организма пресмыкающегося и его соответствия условиям существования.
Существование древних пресмыкающихся с самого начала их возникновения протекало в громадных болотах и болотных лесах, с густой растительностью в прибрежных низменностях и дельтах. С эволюцией наземной растительности, все шире захватывавшей более высокие участки материков, первичные области обитания пресмыкающихся значительно расширились. Однако выход пресмыкающегося из затененных густой растительностью областей на открытые пространства, нагретые тропическим солнцем, был вовсе не так прост, как это нередко кажется исследователям, исходящим в своих представлениях из привычной человеку физиологии организма млекопитающего. Организм пресмыкающегося, не защищенный от перегревания ни специальной термоизоляцией, ни сложным механизмом регулирования постоянной температуры крови, на открытом пространстве днем подвергается опасности. Поэтому древнейшие четвероногие и зверообразные пресмыкающиеся в приспособлениях к обитанию на возвышенных, незатененных участках материка сперва, в конце палеозоя, дали ряд ночных или роющих форм, а еще большее количество, благодаря добавочным приспособлениям в виде фотогормональной терморегуляции с помощью теменного органа, приспособились, к жизни в воде рек, с редкими выходами на сушу. В дальнейшем линия четвероногих пресмыкающихся дала формы с постоянной температурой тела и новыми защитными покровами, иными словами, дала ветвь млекопитающих. Млекопитающие, при значительно более высокой энергии своего организма, в своей приспособительной эволюции смогли обойтись без перехода к двуногому передвижению, т. е. типу движения с наименьшей возможной затратой энергии, столь выгодному для менее активного организма пресмыкающегося. Архозавры обитали в древних условиях наземной жизни — болотах и болотных низменных лесах[, и,] . естественно, что обширное развитие болотных низменностей с густой растительностью в мезозое определило именно развитие архозавров в новых крупных формах — разнообразных динозавров.
Ведущий орган чувств пресмыкающегося — зрение, достигшее весьма совершенного развития. Глаза пресмыкающихся и вообще зауропсид, совершеннее глаз млекопитающих, ведущими чувствами которых являются обоняние и слух. Для ориентировки хищного пресмыкающегося, живущего в местах развития густой растительности, совершенно необходимо иметь высоко поднятую голову и в то же время возможно быстрее передвигаться. Для травоядного, чтобы избегать хищника, необходимы же условия. Оба эти требования основных условий существования разрешаются приспособлением к двуногому передвижению. Поэтому развитие множества двуногих пресмыкающихся есть результат наилучшего приспособления к характерным в древние геологические периоды условиям существования.
Фаунистический состав наиболее крупных местонахождений динозавров в Северной Америке, Европе и Восточной Африке теперь, после изучения условий захоронения динозавров в монгольских местонахождениях, приобретает новое освещение. Становится несомненным зональное распределение преобладающих родов в каждой фауне, что подтверждается и фациальными чертами каждого местонахождения.
Зауроподы — травоядные ящеротазовые динозавры были величайшими наземными животными всех геологических эпох. Их необыкновенные размеры заставляли палеонтологов для объяснения прибегать к самым фантастическим гипотезам. Распространен взгляд, что увеличенная гипофизарная часть мозговой коробки зауропод свидетельствует о патологическом изменении гипофиза, вызвавшем гигантизм.
Водный образ жизни зауропод общепризнан. Однако существуют значительные расхождения в объяснении способа их питания. Маленькая голова и крайне слабое озубление этих животных не дают оснований допустить хищный образ жизни, хотя существуют утверждения о питании зауропод мелкой рыбой, основывающиеся именно на характере озубления.
Если мы попытаемся объяснить строение зауропод, исходя из конкретных условий их существования, то придем к интересным сопоставлениям.
Возникновение зауропод в начале юры и затем их расцвет в юре и начале мела явились ответом на особые существовавшие в этот период условия внешней среды. Середина мезозоя — эпоха весьма низкого стояния материков, сходная с каменноугольным периодом, характерная развитием громадных прибрежных низменностей и лесных болот, с интенсивным углеобразованием. Именно эта часть мезозоя и совпадает с эпохой большой численности зауропод, с их всемирным распространением. Низкие берега материков, очень полого уходившие под уровень моря, образовывали громадную жизненную зону, несомненно, обильную пищей. Однако заселение этой зоны наземными животными долгое время не осуществлялось. Именно для таких берегов характерны огромные приливные волны и очень широкая зона прилива—отлива. Таковы некоторые участки океанских берегов в настоящее время (на Зондских островах, на северо-востоке Австралии, во Флориде, Южной Америке и Африке и т. д.), где низменный берег очень постепенно переходит в морское дно. Ветры и волнения также создают здесь очень сильный накат, резко поднимающий уровень воды.
Гораздо более плоские материки среднего мезозоя были в еще большей степени подвержены приливным волнам. Эти исполинские волны ходили на большом пространстве, сразу поднимая уровень воды в прибрежных лагунах, озерках и болотах на несколько метров, сметая все, что не было приросшим ко дну, силой своего напора. Поэтому древние четвероногие обитатели прибрежий могли жить только в затопленных лесах или на защищенном барьерными рифами берегу. Подобные две зоны захоронения мы видим в карбоне и нижней перми для стегоцефалов и рептилий. Это или: а) каменноугольные леса, или: б) «красные слои» Северной Америки (а также, вероятно Приуралья), непосредственно связанные с рифами.
Эта прибрежная зона, распространившаяся на миллионы квадратных километров в эпоху развития низменностей, пустовала до появления зауропод. Гигантские животные в десятки тонн весом успешно сопротивлялись силе приливных волн, не тонули при подъемах воды и освоили новую, громадную зону обитания, где они не имели конкурентов. В свете высказанных соображений становится понятным развитие у зауропод больших когтей, необходимых для цепляния за грунт.
Серии следов зауропод в Северной Америке найдены во вполне подходящих фациях мелкой лагуны с мягким, но не топким дном.
Приспособительная эволюция зауропод во времени показывает нам картину постепенного освоения все больших глубин прибрежья, возможно, отчасти связанную с постепенным повышением материков и более резким переломом гипсометрической кривой берегов.
Так, если юрские зауроподы могли обитать на глубине до 3 м, верхнеюрские, как диплодок, — до 4—5 м, то нижнемеловые брахиозавры могли кормиться на глубине до 8 м. Развитие зауропод наглядно показывает, как прежний двуногий тип строения скелета (платеозавр) вновь становится четвероногим, но все еще с ведущим значением задних конечностей, более длинных и более тяжелых (диплодок), и, наконец, превращается в жирафоподобный, со значительно сильнее развитыми передними конечностями, покатой назад спиной и сильно поднятой шеей (брахиозавр).
Гигантизм зауропод не был ни случайным, ни закономерным болезненным явлениям, а явился приспособлением к такой обстановке обитания, в которой могли жить только гиганты. Увеличение роста и, вероятно, также ускорение его, для того, чтобы молодые животные быстрее достигали размеров, минимально необходимых для жизни в своей особой обстановке, конечно, требовало усиленного действия гипофиза. Именно тем, что большой гипофиз был нужен для достижения громадного роста, и объясняется увеличение его у зауропод, но вовсе не тем, что увеличение гипофиза (от неизвестных причин) вызвало увеличение роста.
Вопрос о роде питания зауропод пока не может быть разрешен, при недостаточных сведениях о флоре того времени. Однако можно заранее сказать, что челюстной аппарат зауропод так непропорционально мал и слаб по сравнению с их размерами, что растительная пища этих животных должна была быть чрезвычайно мягкой и питательной. Вероятно, в мелких лагунах и болотах середины мезозоя существовала еще неизвестная нам растительность, может быть, типа огромных грибов или питательных студенистых водорослей. Питание рыбой представляется мне невероятным, так как требует и болей сильного озубления и значительно большей подвижности, чем это показывает скелет зауропод с мало разработанными сочленениями. Ловля мелкой рыбешки при помощи громадной шеи с ее полуметровыми позвонками похожа на анекдотическую стрельбу из пушки по воробьям. Такая нелепость не могла существовать в природе. Возможно, что какие-нибудь планктонные животные, икра и личинки могли образовывать в лагунах значительные скопления и служить питательной едой зауропод. Принимая во внимание обилие прибрежных беспозвоночных в мезозое, нужно считать, что этот способ питания зауропод, подобных своего рода четвероногим китам, не заключает в себе ничего невероятного.
Сокращение затопляемых низменных прибрежий с середины мела вызвало сужение ареалов обитания зауропод. Для каждого вида крупных наземных позвоночных площадь обитания должна бить очень большой. Сокращение ее размеров не приводит к простому уменьшению численности животных, но влечет за собой вымирание. Если в середине мезозоя на материковых побережьях лишь сравнительно небольшие участки были недоступны постоянным затоплениям и приливным волнам, то в кайнозое и, особенно. в настоящее время стали редкими, наоборот, пониженные участки. В современных ландшафтах ничто уже не говорит о некогда обширном распространении низменных побережий, вызвавших к жизни чудовищных зауропод.
Область обитания зауропод располагается на самом внешнем крае материка и несколько выдвигается в море. Более высокие области тех же низменных побережий, удаленные от непосредственного соприкосновения с морем, характеризуются зональным распределением различных динозавров (рис. 1).
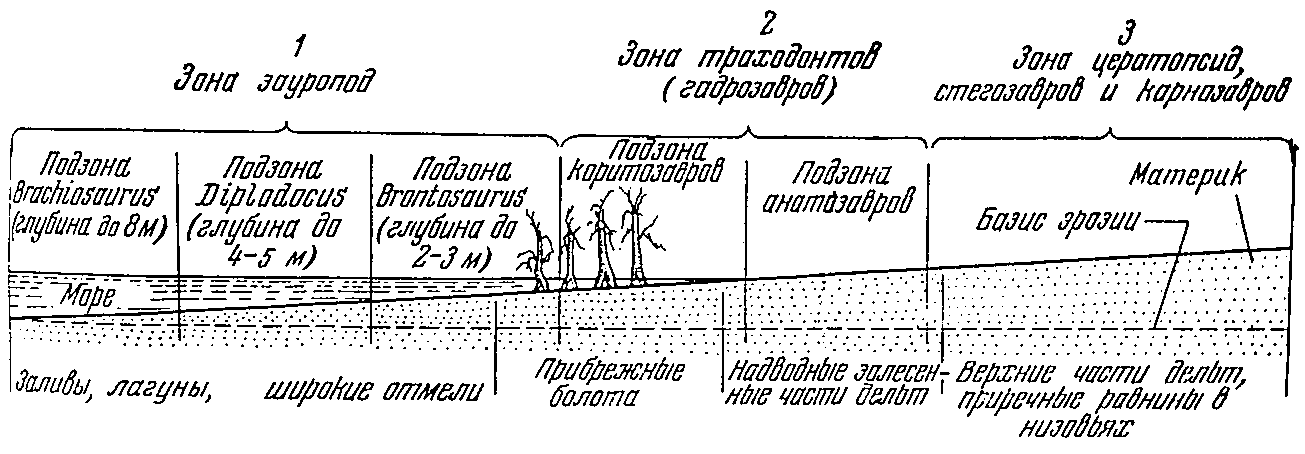
Рис. 1. Схема основных зон обитания динозавров в мезозое. Деревья обозначают границу открытого побережья и низменных прибрежных лесов
В низовьях дельт, защищенных от приливов болотными лесами, в бесчисленных протоках окружающих болот обитали различные виды траходонтных динозавров (гадрозавров), занимавших следующую за зауроподами зону. Зона траходонтов распадается на две подзоны. Одну, с более глубокими водами, занимали коритозавры, паразауролофы и вообще все формы с дополнительными воздушными камерами в носовых полостях и сложным устройством дыхательных путей, с узкими рылами. Вторая, мелководная подзона, служила местом обитания широкорылых анатозавров, в самых общих чертах своей адаптации аналогичных современным бегемотам.
Характерно, что обитатели обеих рассмотренных зон, зауроподы и траходонты, не несут никаких защитных приспособлений как для активной, так и для пассивной защиты. Из этого следует, что обитание в обеих зонах само по себе предохраняло травоядных от непосредственного соприкосновения с хищниками.
Следующая, третья зона, заходящая далеко в материк, захватывает внутреннематериковые окраины прибрежных болотистых лесов и речных низовий и еще не поддается дальнейшему разделению. В этой зоне господствуют хищники — громадные карнозавры, и все обитающие здесь травоядные несут тяжелое панцырное вооружение, появление которого не случайно. В юре и нижнем мелу в третьей зоне находятся стегозавры, в верхнем мелу они заменяются цератопсидами. Здесь же обитают различные анкилозавры от примитивных юрских форм до покрытых тяжелыми костными плитами верхнемеловых сколозавров и анкилозавров.
Распространенные представления о постоянных нападениях карнозавров на зауропод, несомненно, ошибочны. Не говоря уже о том, что среда обитания зауропод была мало доступна, для карнозавров прямое нападение на зауропода, даже вышедшего, скажем, для кладки яиц, в «чужую» зону, могло оказаться успешным лишь в очень редких, исключительных случаях. Траходонты, вероятно, иногда служили пищей карнозаврам, но и тут нападение на крупного траходонта могло окончиться гибелью хищника, так как массивность и сила больших траходонтов были чрезвычайно велики.
Не случайно, что появление наиболее крупных карнозавров — тираннозавров не совпадает со временем наибольшего расцвета зауропод, а случилось позже.
Рассмотренная выше зональность обитания динозавров отражена в большинстве местонахождений. Именно так группируются остатки преобладающих в фаунах родов.
Громадные скопления остатков зауропод в верхней юре и нижнем мелу Уайоминга в Северной Америке или Тендагуру в Восточной Африке отражают захоронение первой, зауроподовой зоны. Остатки хищников или стегозавров там очень редки. Верхнемеловые местонахождения США показывают или ассоциацию траходонтов, или цератопсов с карназаврами. Юрские карнозавры в Европе и Америке встречены вместе со стегозаврами.
Местонахождения Монголии своеобразны — на фоне очень редких остатков зауропод в Нэмэгэту часто встречаются карнозавры и траходонты. Поэтому ассоциацию Нэмэгэту нужно рассматривать как пограничную между зоной траходонтов и зоной карнозавров, со случайно попадавшими сюда зауроподами.
В более низких горизонтах мела (Олгой-Улан-Цаб) чаще встречаются зауроподы, а в местонахождении нижнего мела Хара-Хутул совместно встречаются зауроподы, стегозавры и карнозавры, что, как будто, нарушает это правило. Однако в Хара-Хутул остатки стегозавров и карнозавров встречаются в руслах, промытых между пнями таксодиеобразных мощных деревьев, а зауроподы — с внешней стороны этого ископаемого леса. Таким образом, карнозавры и стегозавры были внесены сюда, в зауроподовую область, из своей, верхней по течению, зоны (рис. 2).
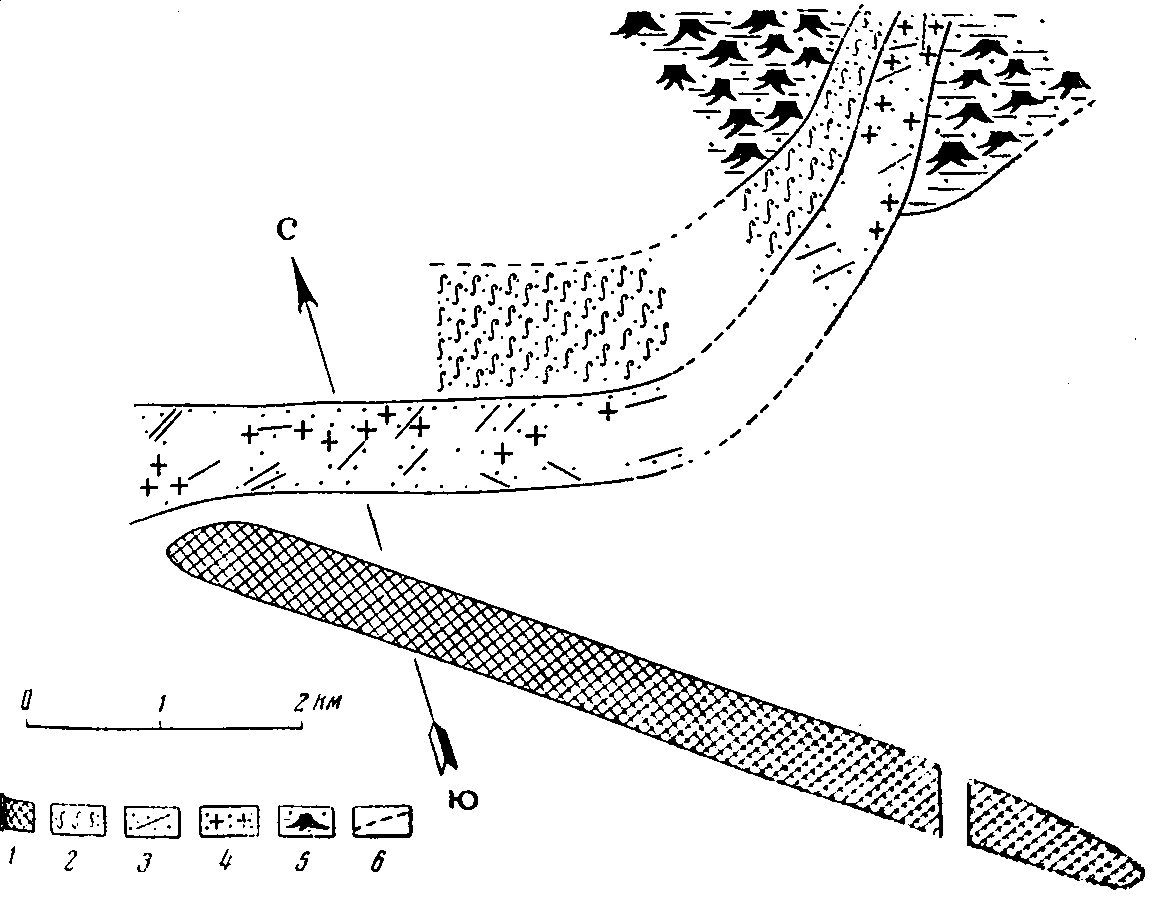
Рис. 2. Схема костоносного русла в нижнемеловых слоях горы Хара-Хутул, Восточная Гоби, МНР.
1 — базальтовая ось хребта; 2 — рыхлые мелкозернистые песчаники с обугленными остатками растения и мелкими стволиками; 3 — грубозернистые песчаники с крупными древесными стволами; 4 — остатки зауропод, стегозавров и хищных динозавров; 5 — слоистые глинистые песчаники с вертикальными пнями таксодиев (?) — область затопленного леса; 6 — предполагаемые границы русла в размытых участках долины.
Огромные русла Нэмэгэту, вмещавшие струи разнообразной скорости и дальности переноса, сделали возможным смешение в захоронении двух разных ассоциаций. Поздние верхние конгломераты Нэмэгэту показывают группировку мелких карнозавров с черепахами, т. е., повидимому обычную ассоциацию лесных форм с речными, встречающуюся в местонахождениях самого различного возраста.
Естественно, что взаимоотношения зон обитания динозавров были сложными и часто на их границах смешивались разные ассоциации. Условия переноса, гибели и вообще вся совокупность процесса захоронения также оказывали серьезное влияние на состав ископаемых фаун. Но тем не менее основная зональность сохраняется и может быть обнаружена по всех месторождениях динозавров. Соответственно с изменениями условий обитания в ходе геологического времени изменяется и преобладание тех или других фаунистических группировок. Для юры и нижнего мела, при наибольшей ширине первой зоны зауропод, наиболее часто в местонахождениях этого времени встречаются зауроподы. В верхнем мелу, при сужении зауроподовой зоны, наиболее часты находки траходонтов, а к концу мела, при наибольшем расширении верхней третьей зоны, чаще всего встречаются цератопсиды, анкилозавры и карнозавры.
Представление о зональном обитании динозавров разрушает установившиеся тенденции к рассмотрению в одном фаунистическом комплексе всех динозавров, постоянно соприкасающихся и конкурирующих, и свидетельствует об ошибочности распространенных реконструкций и картин жизни мезозойских наземных позвоночных.
Первым, поставившим вопрос о значении приливных волн в развитии динозавров, был немецкий ученый Вильфарт (Wilfarth, 1949). В последней своей работе он суммировал и развил свои воззрения. К сожалению Вильфарт впадает в крайность, пытаясь приписать всем или почти всем динозаврам жизнь в зоне приливных волн, вместо того, чтобы признать распространение последних ограниченным, свойственным лишь определенным типам географических ландшафтов прошлого, в которых обитали лишь специально приспособленные типы динозавров.
Вильфарту пришлось допустить повсеместное распространение приливных волн, а для этого придумать ряд совершенно произвольных сопоставлений. Повсеместное распространение приливных волн обязано якобы их гигантской высоте благодаря значительно большей якобы близости луны в мезозойскую эру.
Любое приспособление в морфологических структурах динозавров рассматривается Вильфартом только с точки зрения водного или, скорее, подводного существования. Так, быстро передвигающиеся двуногие динозавры якобы приспособились выпрыгивать со дна на поверхность воды для дыхания, зауроподы заякоривались в илистом грунте кончиком хвоста (!), чтобы противостоять напору огромных волн и т. д. и т. п.
В настоящей статье нет места для подробного критического разбора всех ошибочных и ненаучных допущений Вильфарта. Все они служат лишней иллюстрацией того, как даже вполне правильное и новое по идее научное положение превращается в абсурд при одностороннем его развитии.
Дата добавления: 2020-04-25; просмотров: 194; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!