Капсулы, слизистые слои и чехлы
Снаружи клеточная стенка прокариот часто бывает окружена слизистым веществом. Такие образования в зависимости от структурных особенностей получили название капсул, слизистых слоев или чехлов. Все они являются результатом биосинтеза прокариотами органических полимеров и отложения их вокруг клеток.
Под капсулой понимают слизистое образование, обволакивающее клетку, сохраняющее связь с клеточной стенкой и имеющее аморфное строение (см. рис. 9, 19; 6, 2).Если толщина образования меньше 0,2 мкм и, следовательно, оно может быть обнаружено только с помощью электронного микроскопа, говорят о микрокапсуле. Если больше 0,2 мкм, говорят о макрокапсуле. Последнюю можно видеть в обычный световой микроскоп. Для этого препарат просматривают в капле туши, которая не в состоянии проникнуть в капсулу. На темном фоне выделяются клетки, окруженные светлыми зонами. Если же слизистое вещество имеет аморфный, бесструктурный вид и легко отделяется от поверхности прокариотной клетки, говорят о слизистых слоях, окружающих клетку.
В отличие от капсул чехлы имеют тонкую структуру. Нередко в них обнаруживают несколько слоев с разным строением.Чехлы ряда бактерий, метаболизм которых связан с окислением восстановленных соединений металлов, часто инкрустированы их окислами. Между этими структурами у прокариот обнаружено много переходных форм, так что иногда нельзя четко отграничивать капсулу от слизистых клеточных выделений или капсулу от чехла.
|
|
|
Наличие капсулы зависит от штамма микроорганизма и условий его культивирования. Бактерии, образующие капсулу, могут легко в результате мутации превращаться в бескапсульные формы, что не приводит к какому-либо нарушению клеточной активности, поэтому капсулы нельзя рассматривать как обязательный структурный компонент прокариотной клетки.
Капсулы, слизистые образования и чехлы могут содержать компоненты, одинаковые с клеточной стенкой, однако их химические составы не идентичны. Как правило, химический состав капсул, образуемых бактериями, родо- или видоспецифичен. Основные химические компоненты большинства капсул прокариот – полисахариды гомо- или гетерополимерной природы. Исключение составляет капсула некоторых видов Bacillus , построенная из полипептида, являющегося полимером D-глутаминовой кислоты. Для ряда бактерий показана способность синтезировать и выделять в окружающую среду волокна целлюлозы.
Чехлы как более сложные структуры имеют обычно и более сложный химический состав. Чехол Sphaerotilus natans , например, содержит 36 % cахаров, 11 – гексозамина, 27 – белка, 5,2 – липида и 0,5 % фосфора.
В отличие от капсул чехлы имеют тонкую структуру. Нередко в них обнаруживают несколько слоев с разным строением.Чехлы ряда бактерий, метаболизм которых связан с окислением восстановленных соединений металлов, часто инкрустированы их окислами.Они защищают клетку от механических повреждений, высыхания, создают дополнительный осмотический барьер, служат препятствием для проникновения фагов. Иногда слизистые образования могут служить источником запасных питательных веществ. С помощью слизи осуществляется связь между соседними клетками в колонии, а также прикрепление клеток к различным поверхностям. Способность определенных бактерий синтезировать эти своеобразные внеклеточные полимеры находит практическое применение: их используют в качестве заменителя плазмы крови, а также для получения синтетических пленок.
|
|
|
Жгутики и механизмы движения
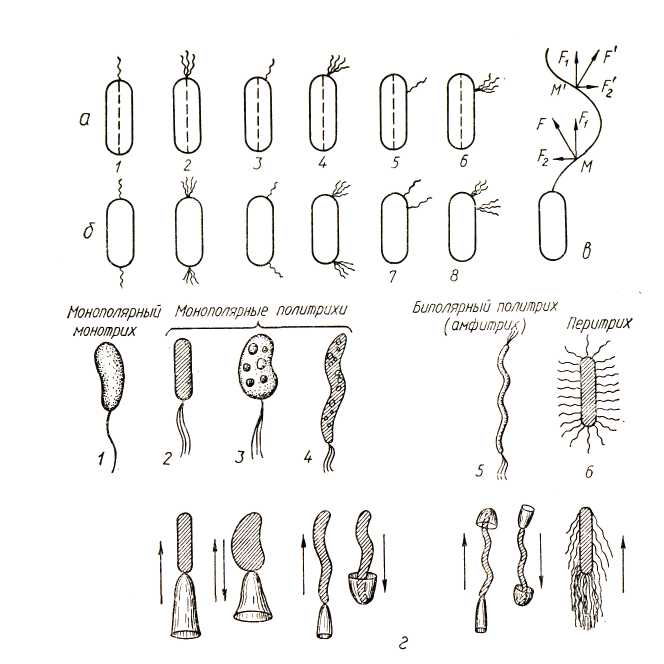
По способности передвигаться все бактерии делятся на две группы – неподвижные и подвижные. Движение бактерий бывает скользящее, вращательное и поступательное с помощью жгутиков. Наиболее распространенный тип движения у бактерий с помощью жгутиков. По количеству и характеру расположения различают жгутики (рис. 14): 1) полюсные, когда один, два и более жгутиков расположены на одном (монополярно) или на обоих (биполярно) полюсах клетки и основание жгутика обычно параллельно длинной оси клетки; 2) подполюсные жгутики (или субполярные), когда один – два и более жгутиков расположены в месте перехода боковой поверхности в полюс клетки на одном или двух концах ее и основание жгутика обычно составляет прямой угол с длинной осью клетки; 3) боковые (или латеральные) жгутики, когда один,два и более жгутиков, в виде кустика, расположены в средней точке одной из половин клетки; 4) перитрихиальные жгутики расположены по всей поверхности клетки по одному или пучками (кишечная палочка, обыкновенный протей), обычно полюса клетки лишены их; 5) смешанные жгутики, когда два или несколько жгутиков расположены в различных точках клетки.
|
|
|
|
|
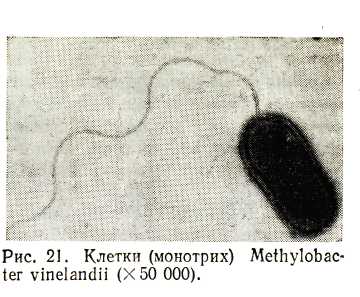
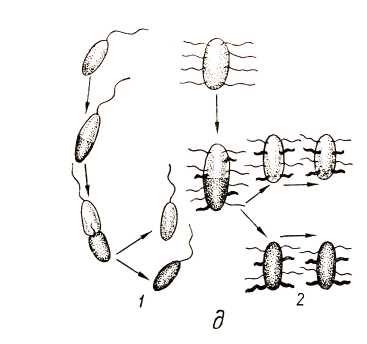
Рис. 14. Жгутики бактерий: жгутики расположены на одном (а) или на обоих (б) концах клетки бактерий: полюсные моно (1) и лофотрихи (2);подполюсные моно (3) и лофотрихи (4); боковые моно- (5) и лофотрихи (в);смешанные жгутики (7, 8); в – схема вращательного и поступательного движения бактерий; г – направление движений бактерий: 1 – Vibrio; 2 – Pseudomonas; 3 – Chromatium; 4 – Thiospirillum; 5 – Spirillum; 6 – Proteus; д – распределение и образование жгутиков во время деления клетки: 1 – клетка с полярным жгутиком; 2 – перитрих
|
|
|
Бактерии имеющие один жгутик, называют монотрихами (рис. 14), пучок жгутиков – лофотрихами. Количество жгутиков у бактерий варьирует в зависимости от вида микроба и условий культивирования, например, у спирилл (лофотрихи) может быть от 5 до 30 жгутиков, у вибрионов – 1 – 2 – 3, у обыкновенного протея, возбудителя столбняка (перитрихи) – 50–1000 жгутиков.
Жгутики очень тонкие образования, их поперечный диаметр колеблется в пределах 10–20 нм у простых жгутиков (белковая нить) до 40–60 нм у сложных жгутиков (когда нить дополнительно одета чехлом, например, у бактерий родов Vibrio, Bdellovibrio, Proteus). Наблюдать жгутики можно темнопольной микроскопией (например, у спирилл), фазово-контрастной микроскопией, с использованием специальных сложных методов окраски с применением протравы (например, танина, который увеличивает диаметр жгутиков и повышает их сродство к краскам), электронной микроскопией. Наиболее ценные данные о строении жгутиков получены при использовании метода электронной микроскопии.
Химический состав жгутиков довольно однообразен и почти полностью представлен белком (98 %), который был назван флагеллином (лат. flagellum – жгутик). Молекулярная масса флагеллина 20–60 тыс. (например, у сенной палочки – 33 000, у кишечной палочки – 60 000), он содержит до 16 аминокислот, среди которых количественно преобладают глутаминовая и аспарагиновая, имеется незначительное количество ароматических аминокислот и отсутствуют триптофан, цистеин, цистин. Флагеллин обладает антигенной специфичностью (его называют Н-антигеном). Четвертичная структура флагеллина представляется полым цилиндром, стенки которого, как предполагают, могут быть «выстланы» глобулярными субъединицами белка под углом к оси жгутика (рис. 15), спирально и в виде прямых тяжей, например, у Salmonella typhimurium. В противоположность жгутикам водорослей и простейших жгутики бактерий не обладают АТФазной активностью.
Обычная толщина жгутика – 10–20 нм, длина – от 3 до 15 мкм. У некоторых бактерий длина жгутика может на порядок превышать диаметр клетки. Как правило, полярные жгутики более толстые, чем перитрихиальные. Жгутик представляет собой относительную жесткую спираль, обычно закрученную против часовой стрелки. Вращение жгутика также осуществляется против часовой стрелки с частотой от 40 до 60 об/с, что вызывает вращение клетки, но в противоположном направлении. Поскольку клетка намного массивнее жгутика, она вращается со значительно меньшей скоростью – порядка 12–14 об/мин. Вращательное движение жгутика преобразуется также в поступательное движение клетки, скорость которого в жидкой среде для разных видов бактерий составляет от 16 до 100 мкм/с.
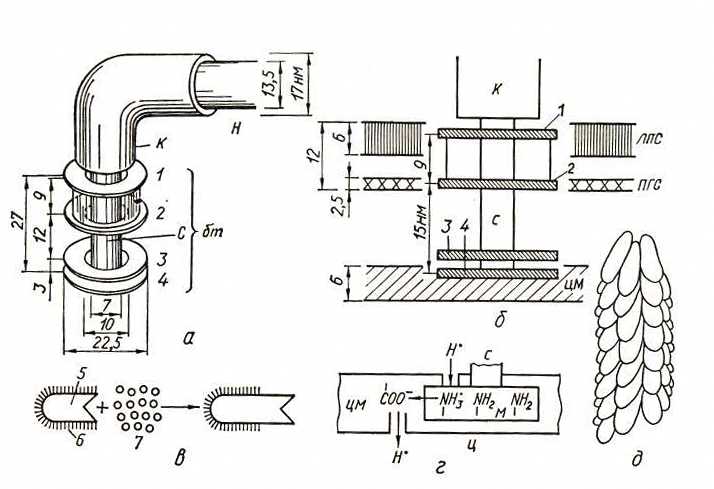
Рис. 15. Строение жгутика бактерий (по Стейниеру Р. и др., 1979; Glagolev A., Sculachev V., 1978; Bode W. и др., 1972). Схема базального конца жгутика (а, б) и его одновекторного роста in vitro (в); модель работы протонного мотора базальной структуры (г); расположение молекул флагеллина, образующих нить жгутика (д). 1, 2 – внешняя пара колец; 3, 4 – внутренняя пара колец; 5 – срезанный фрагмент жгутика, покрытый антителами против жгутиков (6); 7 – субъединицы флагеллина; н – нить; к – крючок; с – стержень; бт – базальное тельце; ц – цитоплазма; цм – цитоплазматическая мембрана; лпс – липополисахаридный слой; пгс – пептидогликановый слой
Крюк (толщина 20–45 нм) состоит из белка, отличающегося от флагеллина, и служит для обеспечения гибкого соединения нити с базальным телом. Базальное тело содержит 9–12 различных белков и представляет собой систему из двух или четырех колец, нанизанных на стержень, являющийся продолжением крюка. Два внутренних кольца – обязательные составные части базального тела, в то время как наружные кольца отсутствуют у грамположительных эубактерий и, следовательно, не необходимы для движения.
Особенности строения базального тела определяются, таким образом, строением клеточной стенки. Интактность последней необходима для движения жгутиковых бактерий. Обработка клеток лизоцимом, приводящая к удалению пептидогликанового слоя клеточной стенки, вызывает и потерю способности бактерий к движению, хотя жгутики остаются при этом неповрежденными.
Изучение строения жгутика под электронным микроскопом обнаружило, что он состоит из трех частей (рис. 16). Основную массу жгутика составляет длинная спиральная нить (фибрилла), у поверхности клеточной стенки переходящая в утолщенную изогнутую структуру – крюк. Нить с помощью крюка прикреплена к базальному телу, вмонтированному в ЦПМ и клеточную стенку.
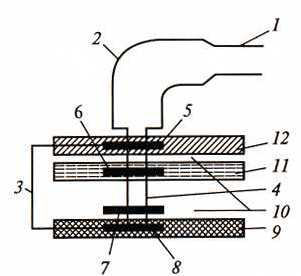
Рис. 16. Строение жгутика грамотрицательных эубактерий: 1 – нить; 2 – крюк; 3 – базальное тело; 4 – стержень; 5 – L-кольцо; 6 – Р-кольцо; 7 – S-кольцо; 8 – М-кольцо; 9 – ЦПМ; 10 – периплазматическое пространство; 11 – пептидогликановый слой; 12– наружная мембрана (по Гусеву М.В. и Минеевой Л.А., 2003)
У большинства прокариот нить состоит только из одного типа белка – флагеллина. Белковые субъединицы уложены в виде спирали, внутри которой проходит полый канал. Наращивание жгутика происходит с дистального конца, куда субъединицы поступают по внутреннему каналу. У некоторых видов жгутик снаружи дополнительно покрыт чехлом особого химического строения или же являющимся продолжением клеточной стенки и, вероятно, построенным из того же материала.
Большие успехи достигнуты в расшифровке механизма движения прокариот, имеющих жгутики. если в клетке много жгутиков, все они при движении собираются в пучок, вращаясь в одном направлении (рис. 17). Вращение жгутиков передается клетке, начинающей вращаться в противоположном направлении, и обеспечивает эффективное движение (плавание) в жидкой среде и более медленное перемещение по поверхности твердых сред.
Для работы двигательного аппарата прокариот необходима энергия. Установлено, что движение жгутиковых прокариот обеспечивается энергией трансмембранного электрохимического потенциала (∆µh+), причем обе его составляющие – электрическая (AT) и концентрационная (АрН) – поддерживают движение. Скорость вращения жгутиков прямо зависит от величины мембранного потенциала. Таким образом, прокариотическая клетка обладает механизмом, позволяющим превращать электрохимическую форму энергии непосредственно в механическую. Молекулярное устройство, обеспечивающее это превращение, к настоящему времени не выяснено, но можно полагать, что оно должно быть весьма эффективным, так как, по проведенным расчетам, энергия, расходуемая на движение, составляет десятые доли процента от общего количества энергетических потребностей клетки.
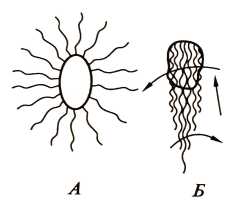
Рис. 17. Клетка Salmonella typhimurium в состоянии покоя (а)и при движении (б).Стрелками показано направление вращения и движения клетки (по Гусеву М.В. и Минеевой Л.А., 2003)
Необычная локализация структур, ответственных за движение, описана у спирохет (рис. 18). Трехслойная структура, окружающая клетку и называемая у спирохет наружным чехлом, аналогична наружной мембране клеточной стенки грамотрицательных бактерий. Этот чехол окружает так называемый протоплазматический цилиндр, состоящий из пептидогликанового слоя клеточной стенки, ЦПМ и цитоплазматического содержимого. Протоплазматический цилиндр обвивается пучком нитчатых структур – аксиальных фибрилл. Число их колеблется от 2 до 100.
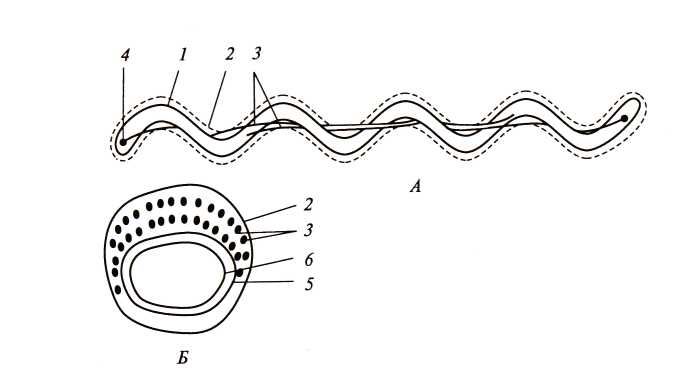
Рис. 18. Клетка спирохеты в продольном (а)и поперечном (б)разрезе. На рис. а изображена клетка, содержащая по одной аксиальной фибрилле у каждого конца; на рис. б – поперечный разрез, прошедший через среднюю часть клетки, где показаны два пересекающихся пучка, состоящих из множества аксиальных фибрилл: 1 – протоплазматический цилиндр; 2 – наружный чехол; 3 – аксиальные фибриллы; 4 – место прикрепления аксиальных фибрилл; 5 – пептидогликановый слой клеточной стенки; 6 – ЦПМ (по Гусеву М.В. и Минеевой Л.А., 2003)
Один конец каждой аксиальной фибриллы прикреплен вблизи полюса протоплазматического цилиндра, другой – свободный. Клетка содержит по два набора фибрилл, прикрепленных субполярно у каждого клеточного конца. Так как каждая аксиальная фибрилла тянется почти вдоль всей длины клетки, пучки фибрилл, прикрепленных у разных полюсов, в центральной части перекрываются.
Изучение строения и химического состава аксиальных фибрилл спирохет обнаружило их близкое сходство с бактериальными жгутиками. Отличие заключается в том, что аксиальные фибриллы спирохет – внутриклеточные структуры, но обеспечивают движение как в жидкой среде, так и по твердому субстрату. Движение спирохет осуществляется за счет вращения фибрилл в периплазматическом пространстве между пептидогликановым слоем и наружной мембраной клеточной стенки, вызывающего эластичную волну на поверхности клеточной стенки. Спирохеты совершают движения трех типов: быстро вращаются вокруг длинной оси спирали, способны к изгибанию клеток и осуществляют передвижение по винтовому или волнообразному пути.
Присущая спирохетам локализация двигательного аппарата интересна тем, что позволяет сделать вывод о возможности его работы в условиях нахождения в «закрытом» клеточными структурами состоянии. Это может служить ключом к пониманию еще одного вида движения, присущего части прокариот, – скольжения. Последнее определяют как способность организма передвигаться по твердому или полужидкому субстрату без помощи наружных локомоторных структур – жгутиков.
Ворсинки
К поверхностным структурам бактериальной клетки относятся также ворсинки и пили).Их насчитывается от нескольких единиц до нескольких тысяч на клетку. Эти структуры не имеют отношения к движению бактерий и обнаружены у подвижных и неподвижных форм.
Строение и функции пилей различны, у одной и той же бактерии могут присутствовать пили разной природы.
Ворсинки (лат. fimbriae – нить, бахрома, волокно) отличаются от жгутиков рядом особенностей: они короче (0,3–1 мкм, редко до 4 мкм), толще, не согнуты волнообразно, встречаются в большом количестве (от 200 до нескольких тысяч), обнаружены у подвижных и неподвижных клеток, не выполняют функции движения. Они кодируются генами хромосомы, состоят из гидрофобного белка – пилина. Ворсинки снижают электрофоретическую подвижность клетки, расположены по всей поверхности клеточной стенки, служат для соединения клеток, защищают клетку от паразитов, препятствуя непосредственному контакту их с поверхностью клетки-хозяина. Некоторые ворсинки кодируются плазмидными генами. Они могут быть одним из факторов патогенности, например, ворсинками клетки кишечной палочки прикрепляются к эпителию. Сократимые полярные пили, возможно, обусловливают дергающее движение бактерий (медленное периодическое перемещение клеток и колоний по поверхности плотной среды), подобное амебоидному движению эукариот.
F -пили присущи мужским клеткам бактерий. Их образование обусловлено наличием полового фактора. Это полые цилиндрические отростки толщиной 8–35 нм, длинной до 1,0–2,0 мкм, образованы белком пилином с молекулярной массой 11 800 (до 40 000). В пилине отсутствуют пролин, цистеин, гистидин, аргинин, имеется много кислых и гидрофобных аминокислот, ковалентно связан с двумя фосфатными группами и остатком D-глюкозы. Пилин синтезируется на рибосомах, связанных с ЦПМ, и накапливается в ней, на ее поверхности осуществляется сборка трубочек пили. Трубочки проходят через клеточную стенку и выступают наружу. Формируются F-пили только у активно растущей клетки за 4–5 мин. и в течение такого же промежутка времени сохраняются на ее поверхности, затем сбрасываются. Половыми ворсинками мужская клетка прикрепляется к женской, образуется конъюгационный тоннель, по которому происходит передача ДНК от донора к реципиенту. У бактерий с половыми ворсинками появляется специфический антиген, чувствительность к мужским мелким РНК- и ДНК-содержащим фагам, клетки становятся малоподвижными, способны к автоагглютинации.
Мембраны
Содержимое клетки отделяется от клеточной стенки цитоплазматической мембраной (ЦПМ) – обязательным структурным элементом любой клетки, нарушение целостности которого приводит к потере клеткой жизнеспособности. На долю ЦПМ приходится 8–15 % сухого вещества клеток. У большинства прокариотных клеток ЦПМ – единственная мембрана. В клетках фототрофных и ряда хемотрофных прокариот содержатся также мембранные структуры, располагающиеся в цитоплазме и получившие название внутрицитоплазматических мембран. Их происхождение и функции будут рассмотрены ниже.
Химический состав мембран. ЦПМ – белково-липидный комплекс, в котором белки составляют 50–75 %, липиды – от 15 до 45 %. Кроме того, в составе мембран обнаружено небольшое количество углеводов. Как правило, липиды и белки составляют 95 % и больше вещества мембран. Главным липидным компонентом бактериальных мембран являются фосфолипиды – производные 3-фосфоглицерина. Хотя у прокариот найдено множество различных фосфолипидов, набор их в значительной степени родо- и даже видоспецифичен. Широко представлены в бактериальных мембранах различные гликолипиды. Стерины отсутствуют у подавляющего большинства прокариот, за исключением представителей группы микоплазм и некоторых бактерий. Из других групп липидов в мембранах прокариот обнаружены каротиноиды, хиноны, углеводороды.
Все липиды эубактерий – производные глицерина – содержат один или несколько остатков жирных кислот, состав которых весьма своеобразен. В основном это насыщенные или мононенасыщенные жирные кислоты с 16–18 углеродными атомами. Поскольку ЦПМ прокариот многофункциональна и участвует в осуществлении разнообразных ферментативных процессов, был сделан вывод, что мембранные белки – это, как правило, ферменты. По аминокислотному составу мембранные белки не отличаются от других клеточных белков, за исключением того, что в них содержится мало (иногда следы) цистеина.
Структура мембран. Мембранные липиды всех эубактерий и части архебактерий образуют бислои, в которых гидрофильные «головы» молекул обращены наружу, а гидрофобные «хвосты» погружены в толщу мембраны (рис. 19). Углеводородные цепи, прилегающие к гидрофильным «головам», довольно жестко фиксированы, а более удаленные части «хвостов» обладают достаточной гибкостью. У некоторых архебактерий (ряд метаногенов, термоацидофилы) мембранные липиды, в состав которых входит С40-спирт, формируют монослойную мембрану, по толщине равную бислойной. Монослойные липидные мембраны обладают большей жесткостью сравнительно с бислойной. При «биологических» температурах мембранные липиды находятся в жидкостно-кристаллическом состоянии, характеризующемся частичной упорядоченностью структуры. При понижении температуры они переходят в квазикристаллическое состояние.
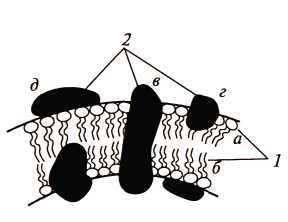
Рис. 19. Модель строения элементарной биологической мембраны: 1 – молекулы липидов: а – гидрофильная «голова»; б – гидрофобный «хвост»; 2 – молекулы белков: в – интегральная; г – периферическая; д – поверхностная (по Гусеву М.В. и Минеевой Л.А., 2003)
В отличие от липидов у мембранных белков нет единого способа структурной организации. 30–50 % белка имеет конфигурацию α-спирали, остальная часть находится преимущественно в виде беспорядочного клубка. Вероятно, часть белков лишена ферментативной активности и участвует только в поддержании мембранной структуры. В то же время доказано, что для осуществления белками некоторых функций необходима их строго упорядоченная взаимная организация в мембране.
Предложено несколько моделей строения мембраны. Наибольшее признание получила модель, учитывающая большинство данных, известных о мембранах, согласно которой в липидную основу включены асимметрично расположенные белковые молекулы (см. рис. 19). Некоторые из них образуют скопления на поверхностях липидного би- или монослоя, другие частично или полностью погружены в него, третьи пронизывают его насквозь. В модели подчеркнута асимметрия строения мембраны, основанная на различиях в химическом строении и расположении молекул белка.
Функции ЦПМ прокариот. ЦПМ прокариот выполняет разнообразные функции, в основном обеспечиваемые локализованными в ней соответствующими ферментными белками. Первоначально была постулирована барьерная функция клеточной мембраны, получившая позднее экспериментальное подтверждение. С помощью специальных переносчиков, называемых транслоказами, через мембрану осуществляется избирательный перенос различных органических и неорганических молекул и ионов. В ней локализованы ферменты, катализирующие конечные этапы синтеза мембранных липидов, компонентов клеточной стенки и некоторых других веществ.
Общепризнана роль ЦПМ прокариот в превращениях клеточной энергии. У бактерий, источником энергии для которых служат процессы дыхания или фотосинтеза, в ЦПМ определенным образом расположены переносчики цепи электронного транспорта, функционирование которых приводит к генерированию электрохимической энергии (∆µН+), используемой затем в клетке по разным каналам, в том числе и для образования химической энергии (АТФ). ЦПМ является одним из компонентов аппарата генерирования ∆µН+. В мембране расположены также ферментные комплексы, обеспечивающие превращения: ∆µН+ ←→АТФ. ЦПМ принимает участие в репликации и последующем разделении хромосомы прокариотной клетки.
В последнее время выявляется еще одна функциональная грань клеточных мембран – их интегрирующая роль в организме, вполне сочетающаяся с давно установленной разъединяющей (барьерной) функцией. Клетка – единое целое. В обеспечении этого принципа клеточной организации важная роль принадлежит мембранам. Показан перенос электрохимической энергии и электронов вдоль мембран. Последние рассматриваются так же как возможные пути транспорта жирорастворимых субстратов и молекулярного кислорода.
ЦПМ является основным барьером, обеспечивающим избирательное поступление в клетку и выход из нее разнообразных веществ и ионов. У грамположительных форм ЦПМ является и единственным барьером такого рода, у грамотрицательных эубактерий функции дополнительного барьера (молекулярного «сита») выполняет наружная мембрана клеточной стенки, через которую молекулы транспортируются только по механизму пассивной диффузии.
Большинство (если не все) гидрофильных веществ поступает в клетку за счет функционирования систем, в состав которых входят специальные переносчики (транслоказы, или пермеазы), так как скорость физической диффузии этих веществ через гидрофобный слой мембраны очень невелика. Переносчики – вещества белковой природы, локализованные в мембране и характеризующиеся высокой субстратной специфичностью, – связываясь с субстратом, подвергаются конформационным изменениям и вследствие этого приобретают способность к перемещению субстрата с одной стороны ЦПМ на другую.
Известен механизм транспорта, получивший название облегченной диффузии, требующий для переноса веществ через мембрану участия транслоказ. Перенос веществ в этом случае происходит по градиенту их концентрации и не требует энергетических затрат. Этот механизм транспорта не получил широкого распространения у прокариот. Основным механизмом избирательного переноса веществ через ЦПМ прокариот является активный транспорт, позволяющий «накачивать» в клетку молекулы и ионы против их концентрационных и электрических градиентов. Этот процесс, так же как и облегченная диффузия, протекает при участии локализованных в ЦПМ переносчиков белковой природы с высокой специфичностью к субстрату, но в отличие от облегченной диффузии для движения против электрохимического градиента требует затрат метаболической энергии. Транспорт такого рода должен быть поэтому сопряжен с реакциями, продуцирующими энергию в химической или электрохимической форме.
Во всех описанных выше путях переноса веществ через ЦПМ они поступают в клетку в химически неизмененном виде. У прокариот известны системы транспорта, с помощью которых осуществляется поступление в клетку ряда сахаров, при этом процесс их переноса через мембрану сопровождается химической модификацией молекул. Так происходит, например, поступление в клетки многих прокариот молекул глюкозы, в процессе которого они фосфорилируются.
Внутрицитоплазматические мембраны прокариот
Среди внутрицитоплазматических мембран выделяют несколько видов. Развитая система внутрицитоплазматических мембран характерна для большинства фотосинтезирующих эубактерий. Поскольку было показано, что в этих мембранах локализован фотосинтетический аппарат клетки, они получили общее название фотосинтетических мембран.
У прокариот, принадлежащих к разным группам, описаны локальные впячивания ЦПМ, получившие название мезосом (см. рис. 9).
Эти внутриклеточные мембранные образования впервые были описаны у Bacillus cereus. По морфологическим особенностям различают ламеллярные (пластинчатые), везикулярные (имеющие форму пузырьков), тубулярные (трубчатые) мезосомы (рис. 20). Часто в бактериальной клетке наблюдаются мезосомы смешанного типа, состоящие из ламелл, трубочек, пузырьков, мезосомные комплексы. Мезосомный комплекс (рис. 20) ограничен инвагинацией ЦПМ мешковидной формы, содержит ветвящиеся внутренние трубочки и пластинчатые мембраны.
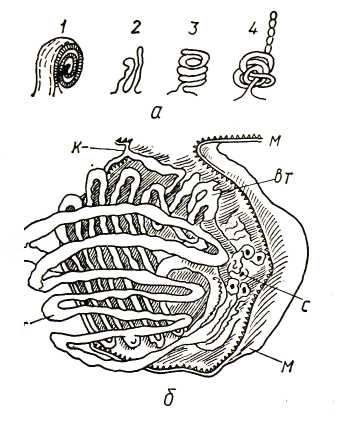
Рис. 20. Типы истинных мезосом (а) и строение мезосомного комплекса (б) (по Бирюзовой В.И., Поглазовой М.Н., 1977; Burdett I., 1972): 1 – ламеллярный; 2–4 – тубулярный типы; I – ветвящиеся внутренние трубочки: к – контакт внутренних мембран с мембраной мешочка; м – мембрана, окружающая мешочек; с – скопление пластинчатых мембранных элементов
Хорошо развитые и сложно организованные мезосомы характерны для грамположительных эубактерий. У грамотрицательных видов они встречаются значительно реже и относительно просто организованы. По расположению в клетке различают мезосомы, образующиеся в зоне клеточного деления и формирования поперечной перегородки (септы), мезосомы, к которым прикреплен нуклеоид, и мезосомы, сформированные в результате инвагинации периферических участков ЦПМ.
Существуют разные точки зрения относительно роли мезосом в клетке. Согласно одной из них мезосомы не являются обязательной структурой, а служат только для усиления определенных клеточных функций, увеличивая общую «рабочую» поверхность мембран. Получены данные о том, что с мезосомами связано усиление энергетического метаболизма клеток. Мезосомы играют роль в репликации хромосомы и ее последующем расхождении по дочерним клеткам, участвуют в процессе инициации и формирования поперечной перегородки при клеточном делении. Для некоторых грамположительных бактерий обнаружено участие мезосом в секреторных процессах.
Высказывается также предположение, что мезосомы не принимают активного участия в процессах клеточного метаболизма, но выполняют структурную функцию, обеспечивая компартментализацию прокариотной клетки, т. е. пространственное разграничение внутриклеточного содержимого на относительно обособленные отсеки, что создает более благоприятные условия для протекания определенных последовательностей ферментативных реакций.
Цитоплазма и рибосомы
Содержимое клетки, окруженное ЦПМ, называется цитоплазмой. Фракция цитоплазмы, имеющая гомогенную консистенцию и содержащая набор растворимых РНК, ферментных белков, продуктов и субстратов метаболических реакций, получила название цитозоля. Другая часть цитоплазмы представлена разнообразными структурными элементами: внутрицитоплазматическими мембранами (если они есть), генетическим аппаратом, рибосомами и включениями разной химической природы и функционального назначения.
Рибосомы – место синтеза белка – рибонуклеопротеиновые частицы размером 15///– 20 нм. Их количество в клетке зависит от интенсивности процессов белкового синтеза и колеблется от 5000 до 90 000. Общая масса рибосом может составлять примерно 1/4 клеточной массы, а количество рибосомальной РНК (рРНК) – 80///85 % всей бактериальной РНК. Отношение рРНК/ белок в рибосомах Е. coli составляет 2 : 1, у других прокариот оно может быть несколько сдвинуто в сторону преобладания белка. Рибосомы прокариот имеют константу седиментации 70S[**], отчего получили название 70S-частиц. Они построены из двух неодинаковых субчастиц: 30S- и 50S-субъединиц. 30S-частица содержит одну молекулу 16S-рРНК и в большинстве случаев по одной молекуле белка более 20 видов. 50S-субъединица состоит из двух молекул рРНК (23S и 5S).В ее состав входят более 30 различных белков, также представленных, как правило, одной копией. Большая часть рибосомальных белков выполняет структурную функцию.
Синтез белка осуществляется агрегатами, состоящими из рибосом, молекул информационной и транспортных РНК и называемыми полирибосомами, или полисомами. Последние могут находиться в цитоплазме или же быть связанными с мембранными структурами.
Дата добавления: 2019-09-13; просмотров: 375; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!