Структурные и генетические отличия прокариот и эукариот
| А. Структурной | ||
| 1. Замкнутые вторичные полости, создаваемые мембранами | Отсутствуют | Имеются |
| 2. Эндоплазматический ретикулум | Отсутствует | Имеется |
| 3. Размер цитоплазматических рибосом | 70S | 80S |
| 4. Наличие рибосом в органеллах | Отсутствуют | В митохондриях (70S) |
| 5. Часть рибосом ассоциирована с цитоплазматической мембраной | Наблюдается | Отсутствует |
| 6. Лизосомы | Отсутствуют | Имеются |
| 7. Митохондрии | Отсутствуют | Имеются |
| 8. Органеллы, окруженные белковой однослойной мембраной | Имеются | Отсутствуют |
| 9. Хлоропласта | Отсутствуют | Имеются или отсутствуют |
| 10. Аппарат Гольджи | Отсутствует | Имеется |
| 11. Ядро | Имеется аналог ядра-нуклеоид | Имеется ядро |
| 12. Ядерная мембрана | Отсутствует | Имеется |
| 13. Ядрышко | Отсутствует | Имеется |
| 14. Вакуоли | Встречаются редко | Встречаются часто |
| 15. Жгутики | Состоят из одной или нескольких фибрилл | Каждый жгутик состоит из 20 фибрилл, собранных в группы 2×9+2 |
| Особенности организации | Прокариоты | Эукариоты | ||||
| Б. Генетической |
| |||||
| 1. Организация ядерной ДНК | ДНК не отделена от цитоплазмы мембраной | Ядерная оболочка окружена мембраной | ||||
| 2. Хромосома | Одна кольцевая (в клетке несколько ее копий) | Больше одной | ||||
| 3. Гистоны в хромосоме | Отсутствуют (у некоторых прокариот обнаружены гистоноподобные белки) | Имеются | ||||
| 4. Хромосома представляет собой репликон
| Единый репликон | Множество репликонов | ||||
| 5. Интронэкзонная структура хромосомы и сплайсинг считываемых РНК | Отсутствует | Имеется | ||||
| 6. Гены организованы в опероны | Имеются | Обычно отсутствуют | ||||
| 8. Для инициации белкового синтеза присутствие на 5’-конце иРНК | Не обязательно | Необходимо | ||||
| 9. Митоз | Отсутствует | Имеется | ||||
| 10. Мейоз | Отсутствует | Имеется | ||||
| 11. Концентрация ДНК | В нуклеоиде и цитоплазматической ДНК, не окруженной оболочкой | В ядре, окруженном оболочкой | ||||
| 13. Способы генетической рекомбинации | Отсутствует | Имеется | ||||
| 14.Образование частичных диплоидов при однонаправленном переносе ДНК | Имеется | Отсутствует | ||||
|
2. Функциональные и химические различия прокариот и эукариот | ||||||
| Отличия | Прокариоты | Эукариоты | ||||
| А. Функциональные |
|
| ||||
| 1. Дыхательная система | Является частью мембран или мезосом | Осуществляется в митохондриях – мембранных органеллах | ||||
| Отличие | Прокариоты | Эукариоты |
| 2. Движение цитоплазмы | Отсутствует | Часто обнаруживается |
| 3. Анаэробиоз | Факультативный и облигатный | Факультативный |
| 4. Фагоцитоз | Отсутствует | Имеется |
| 5. Пиноцитоз | Отсутствует | Имеется |
| 6. Внутриклеточное пищеварение | Отсутствует | Имеется |
| 7. Устойчивость к облучению | Очень высокая | Низкая |
| 8. Верхний предел температуры | 75–90 °С | 40–60 °С |
| 9. Секреция веществ в пузырьках Гольджи | Отсутствует | Имеется |
| 10. Клеточные эндосимбионты | Отсутствуют | Имеются |
| Б. Химические | ||
| 1.Пептидогликаны в составе клеточной стенки | Имеются | Отсутствуют |
| 2. Тейхоевые кислоты в составе клеточной стенки | Имеются | Отсутствуют |
| 3. Стерины в составе мембран | Отсутствуют (некоторые микоплазмы включают их в мембрану из питательной среды) | Имеются |
| 4. ДНК в органеллах | Отсутствует | Имеется |
Такие мембраны называются элементарными. Обязательными химическими компонентами каждой клетки являются два вида нуклеиновых кислот (ДНК и РНК), белки, липиды, углеводы. Цитоплазма и элементарная мембрана, окружающая ее, – непременные и обязательные структурные элементы клетки. Это то, что лежит в основе строения всех без исключения клеток. Изучение тонкой структуры выявило существенные различия в строении клеток прокариот (бактерий и цианобактерий) и эукариот (остальные макро- и микроорганизмы).
|
|
|
Прокариотическая клетка отличается тем, что имеет одну внутреннюю полость, образуемую элементарной мембраной, называемой клеточной, или цитоплазматической (ЦПМ). У подавляющего большинства прокариот ЦПМ – единственная мембрана, обнаруживаемая в клетке. В эукариотных клетках в отличие от прокариотных есть вторичные полости. Ядерная мембрана, отграничивающая ДНК от остальной цитоплазмы, формирует вторичную полость. Наружные мембраны хлоропластов и митохондрий, окружающие заключенные в них функционально специализированные мембраны, играют аналогичную роль. Клеточные структуры, ограниченные элементарными мембранами и выполняющие в клетке определенные функции, получили название органелл. Ядро, митохондрии, хлоропласта – это клеточные органеллы. В эукариотных клетках помимо перечисленных выше есть и другие органеллы.
|
|
|
В клетках прокариот органеллы, типичные для эукариот, отсутствуют. Ядерная ДНК у них не отделена от цитоплазмы мембраной. В цитоплазме находятся функционально специализированные структуры, но они не изолированы от цитоплазмы с помощью мембран и, следовательно, не образуют замкнутых полостей. Эти структуры могут быть сформированы и мембранами, но последние не замкнуты и, как правило, обнаруживают тесную связь с ЦПМ, являясь результатом ее локального внутриклеточного разрастания. В клетках прокариот есть также образования, окруженные особой мембраной, имеющей иное по сравнению с элементарной строение и химический состав.
|
|
|
В связи с тем что прокариотическая и эукариотная организация клеток принципиально различны, было предложено только н основании этого признака выделить все прокариоты в особое царство. Р. Меррей (R. Murray) в 1968 г. предложил все клеточные организмы разделить на две группы по типу их клеточной организации: царство Prokaryotae, куда вошли все организмы с прокариотным строением клетки, и царство Eukaryotae, куда включены все высшие протесты, растения и животные.
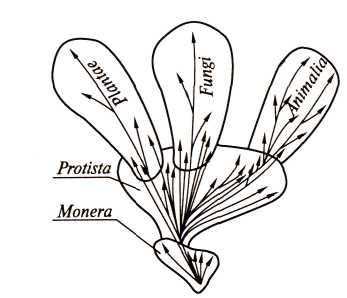
Рис. 6. Схема пяти царств живого мира: прокариоты (царство Мопега), одноклеточные эукариоты (царство Protista), многоклеточные эукариоты (царства Plantae, Fungi, Animalia) (по Whittaker, 1969)
Р. Виттэкер (R.Whittaker) предложил схему, по которой все живые организмы, имеющие клеточное строение, представлены разделенными на пять царств (рис. 6). Такая система классификации живого мира отражает три основных уровня его клеточной организации: Мопега включает прокариотные организмы, находящиеся на самом примитивном уровне клеточной организации; Protista – микроскопические, в большинстве своем одноклеточные, недифференцированные формы жизни, сформировавшиеся в результате качественного скачка в процессе эволюции, приведшего к возникновению эукариотных клеток; многоклеточные эукариоты представлены, в свою очередь, тремя царствами Plantae, Fungi и Animalia.
Три последние таксономические группы различаются по способу питания: фототрофный тип питания за счет процесса фотосинтеза характерен для растений (Plantae); грибы (Fungi) в основном характеризуются осмотрофным типом питания, т. е. питанием растворенными органическими веществами; животные (Animalia) осуществляют голозойное питание, заключающееся в захватывании и переваривании твердой пищи. Способы питания, специфические для растений и грибов, возникли в процессе эволюции на уровне Мопега. На уровне Protista они получили свое дальнейшее развитие; здесь же сформировался третий тип питания – голозойный.
Можно с определенностью утверждать, что обособление прокариотных микроорганизмов в отдельное царство Prokaryotae правомерно, поскольку основано на принципиальных различиях в структуре прокариотных и эукариотных клеток, т. е. тех единиц, из которых построены все клеточные формы жизни.
Глава 2. СИСТЕМАТИКА ПРОКАРИОТ.
ГРУППЫ ПРОКАРИОТНЫХ ОРГАНИЗМОВ
Систематика (таксономия) – наука о многообразии и взаимосвязях между организмами. Одна из задач систематики – распределение (классификация) множества организмов по группам (таксонам)[§]. Но прежде чем осуществлять такое распределение, необходимо достаточно полно охарактеризовать объекты и на основании отобранной информации идентифицировать их. Последнее может привести к выявлению организмов с неизвестными или известными признаками и соответственно помещению их в новый таксон на определенном уровне или же отнесению к известным таксонам.
Для характеристики организмов используют разнообразные признаки: морфологические, цитологические, культуральные, физиологические, биохимические, иммунологические и др. Если объем информации для характеристики объектов по существу беспределен, как бесконечен сам процесс познания природы, то для целей идентификации может быть использован ограниченный объем информации, достаточный для распределения организмов по таксономическим группам.
Специальный раздел таксономии – номенклатура – имеет дело с правилами присвоения наименований описанным объектам. В систематике бактерий для наименования объекта используют биномиальную номенклатуру К. Линнея (К. Linne, 1707–1778), согласно которой биологическому виду присваивают название, состоящее из двух слов: первое определяет принадлежность организма к определенному роду, второе – виду. Названия бактериям присваивают в соответствии с правилами Международного кодекса номенклатуры бактерий.
Основной таксономической категорией является вид. По современным представлениям, вид – это группа близких между собой организмов, имеющих общий корень происхождения и на данном этапе эволюции характеризующихся определенными морфологическими, биохимическими и физиологическими признаками, обособленных отбором от других видов и приспособленных к определенной среде обитания.
Важным признаком, определяющим принадлежность организмов к одному виду, является их способность скрещиваться и давать жизнеспособное потомство. Однако у прокариот размножение половым путем отсутствует, поэтому данный признак для определения видовой принадлежности к ним неприменим. Отнесение прокариотных организмов к одному или разным видам осуществляется в большой степени эмпирическим путем на основе анализа многих признаков, при этом генетическая информация, содержащаяся в нехромосомных генетических элементах, для определения видовой принадлежности не используется.
Виды объединяют в таксоны более высокого порядка – роды, роды – в семейства, далее следуют порядки, классы, отделы, царства. Для высших таксономических категорий пока нет удовлетворительного определения.
В микробиологии употребляются такие термины, как «штамм» и «клон». Под штаммом понимают бактериальные культуры одного вида, выделенные, например, из разных мест обитания. Различия между штаммами не выходят за пределы вида. Клон – еще более узкое понятие, это культура, выделенная из одной клетки.
Существуют 2 типа систематики биологических объектов: филогенетическая, или естественная, в основе которой лежит установление родственных (генетических, эволюционных) связей между организмами, и практическая, или искусственная, цель которой – выявление степени сходства между организмами для быстрой их идентификации и установления принадлежности к определенным таксонам. Если существующая систематика высших организмов отражает в определенной мере эволюционные связи между ними, т. е. признаки, используемые для выявления степени сходства, отражают и степень родства между этими организмами, то попытка создания на этой же основе систематики прокариот не была успешной.
Важным шагом в развитии систематики прокариот явилось использование признаков, дающих информацию о химическом строении клетки: состав оснований ДНК, ДНК–ДНК – и ДНК – РНК-гомологии, аминокислотная последовательность белков, строение рибосом, компонентов клеточной стенки и т. д.
Наиболее полно задача быстрой идентификации прокариотных организмов решается с помощью Определителя бактерий Берги, выпускаемого периодически Обществом американских бактериологов с привлечением крупных специалистов в области изучения тех или иных групп бактерий. Первое издание Определителя было выпущено в 1923 г. группой американских бактериологов под руководством Д.X. Берги (D.H. Bergey, 1860–1937); девятое издание в 4 томах вышло в 1984–1989 гг.
В девятом издании Определителя бактерий Берги все обнаруженные организмы, отнесенные в царство Prokaryotae, разделены на 33 группы. Признаки, по которым осуществляется разделение на группы, как правило, относятся к категории легко определяемых и вынесены в названия групп, например: грамотрицательные аэробные палочки и кокки (группа 4), анаэробные грамотрицательные кокки (группа 8), грамположительные палочки и кокки, образующие эндоспоры (группа 13), скользящие бактерии, образующие плодовые тела (группа 24). Основная идея классификации «по Берги» – легкость идентификации бактерий. Для осуществления этого используют совокупность признаков: морфологических (форма тела; наличие или отсутствие жгутиков, капсулы; способность к спорообразованию; особенности внутриклеточного строения; окрашивание по Граму), культуральных (признаки, выявляемые при культивировании в лаборатории чистой культуры), физиолого – биохимических (способы получения энергии; потребности в питательных веществах; отношение к факторам внешней среды; нуклеотидный состав и последовательность нуклеотидов в молекуле ДНК; наличие и характер минорных оснований в ДНК; нуклеотидный состав рибосомальной РНК; последовательность аминокислот в ферментных белках с аналогичными функциями).
Ценность Определителя в том, что он представляет собой наиболее полную сводку известных бактериальных форм и самое современное пособие для идентификации бактерий.
Представленная в Определителе бактерий Берги система классификации является строго идентификационной и не решает задачи выявления эволюционных связей между прокариотами. В то же время конечной целью является построение такой системы, в основе которой лежали бы родственные связи между прокариотными организмами.
Важный шаг на пути создания естественной систематики прокариот связан с успехами молекулярной биологии. В 60-х гг. XX в. было установлено, что все свойства организма определяются уникальными химическими молекулами – ДНК, поэтому бактерии могут быть классифицированы путем сравнения их геномов. По такому признаку, как генетический материал, оказалось возможным на основании выявления степени сходства делать вывод о степени родства между организмами. Первоначально для таксономических целей сравнивали молярное содержание суммы гуанина и цитозина (ГЦ) в процентах от общего количества оснований ДНК у разных объектов. Этот показатель у прокариот колеблется от 25 до 75 %. Однако ГЦ-показатель дает возможность только для грубого сравнения геномов. Если организмы имеют одинаковый нуклеотидный состав ДНК, возможно и сходство, и различие между ними, поскольку генетическое кодирование основано не только на определенном содержании оснований в единице кодирования (триплете), но и на их взаимном расположении.
Более тонкий метод оценки генетического сходства организмов – сравнение нуклеотидных последовательностей ДНК из разных источников методом ДНК–ДНК-гибридизации. Метод наиболее полезен для классификации на уровне вида, т. е. в случае высокой степени гомологии, и малоинформативен для классификации объектов на уровне высоких таксонов. В то же время часто несовпадение выводов, сделанных на основании фенотипических признаков и ДНК-гибридизации. В целом значение данных о строении ДНК для систематики прокариот огромно, так как позволяет перейти от установления степени сходства к выводам о степени родства между организмами.
Колебания нуклеотидного состава ДНК у эукариотных микроорганизмов (молярная доля, %): грибы – 26–70, водоросли – 37–68, простейшие – 22–68; у высших растений и животных – 35–45. Колебания в составе оснований ДНК вирусов приблизительно такие же, как у прокариот.
Все прокариотические микроорганизмы объединены в царство Procaryotae, которое подразделяется на четыре отдела. Они, в свою очередь, делятся на секции, классы, порядки, семейства, роды, виды.
Отдел I . Gracilicutes (от лат. gracilus – тонкий, стройный, cutes – кожа). Включает в себя грамотрицательные микроорганизмы. В отделе девять секций.
Секция 1. Спирохеты. Порядок Spirochaetales. Включает в себя два семейства: Spirochaetaceae (четыре рода), Leptospiraceae (один род).
Секция 2. Спиралевидные и изогнутые аэробы (микроаэрофилы). Одно семейство – Spirillaceae, в котором шесть родов. Патогенны для человека и животных микроорганизмы рода Campylobacter .
Секция 3. Грамотрицательные неподвижные изогнутые бактерии. Одно семейство – Spirosomonaceae, в котором патогенных три рода.
Секция 4. Аэробные грамотрицательные палочки, округлые и кокки. Восемь семейств, два из которых имеют патогенные микроорганизмы. Семейство Pseudomonadaceae включает в себя четыре рода, более 25 видов, среди которых имеются патогенные ( P . mallei и др.). Семейство Neisseriaceae имеет 16 родов. Роды Neisseria и Moraxella содержат патогенные для человека и животных микроорганизмы. Роды Bordetella , Brucella и Francisella не внесены в семейства: содержат патогенные для человека и животных виды.
Секция 5. Грамотрицательные факультативные анаэробы. Три семейства: Enterobacteriaceae, Vibrionaceae и Pasteurellaceae. Семейство Enterobacteriaceae имеет 14 родов (Escherichia, Salmonella, Citrobacter, Klebsiella, Enterobacter, Erwinia, Shigella, Proteus, Yersinia и др.). Семейство Vibrionaceae имеет два рода. В род Vibrio включены патогенные микроорганизмы. Семейство Pasteurellaceae имеет три основных рода: Pasteurella , Haemophilus и Actinobacillus . Содержат патогенные виды микроорганизмов.
Секция 6. Строгие анаэробы. Изогнутые грамотрицательные палочки. Одно семейство – Bacteroidaceae, в котором 13 родов, среди которых имеются патогенные.
Секция 7. Диссимилирующие и разлагающие сульфат бактерии. Семь непатогенных родов.
Секция 8. Анаэробные грамотрицательные кокки. Одно семейство – Vellonellaceae, в котором три рода.
Секция 9. Риккетсии и хламидии. Два порядка: Rickettsiales и Chlamydiales. Порядок Rickettsiales имеет три семейства: Rickettsiaceae, Bartonellaceae и Anaplasmataceae. Семейство Rickettsiaceae имеет три трибы, в которые внесено восемь родов. Семейство Bartonellaceae содержит два рода, a Anaplasmataceae – четыре. Порядок Chlamydiales имеет одно семейство Chlamydiaceae и один род – Chlamydia . Все семейства содержат патогенные микроорганизмы.
Отдел II. Firmicutes (от лат. firmis – крепкий, cutes – кожа). В отдел включены главным образом грамположительные бактерии.
Секция 12. Грамположительные кокки. Два семейства: Micrococcaceae и Deinococcaceae. Семейство Micrococcaceae имеет четыре рода: Micrococcus , Stomatococcus , Planococcus , Staphylococcus .
В секцию кроме указанных двух семейств внесены десять самостоятельных родов: Streptococcus , Leuconostos , Pedicoccus , Sarcina и др.
Секция 13. Спорообразующие грамположительные палочки и кокки. Шесть родов: Bacillus, Clostridium, Sporolactobacillus, Sporosarcina и др. Первые два рода имеют патогенные виды.
Секция 14. Неспорообразующие грамположительные палочки. Семь родов: Lactobacillus, Listeria, Erysipelotrix и др. Имеются патогенные.
Секция 15. Неспорообразующие внутриклеточные грамположительные палочки. 21 род: Corynebacterium, Mi с robacterium, Propionibacterium, Eubacterium, Asotobacterium, Bifidobacterium, Actinomices и др.
Секция 16. Микобактерии. Одно семейство Mycobacterioсеае. Семейство имеет один род Mycobacterium, в котором 49 видов: М. tuberculosis , М. bovis , М. avium , М. paratuberculosis , М. leprae и др.
Секция 17. Nocardioforms. Девять родов: Nocardia , Pseudococcus , Pseudonocardia и др.
Отдел III . Tenericutes. Объединены грамотрицательные прокариоты без клеточной стенки, но имеющие цитоплазматическую мембрану. В отделе десятая секция – микоплазмы, класса Mollicutes (от лат. molli – мягкий, cutes – покров, кожа). В классе один порядок – Mycoplasmatales – и три семейства: Mycoplasmataceae, Acholeplasmataceae, Spiroplasmataceae. В основном патогенные микоплазмы включены в семейство Mycoplasmataceae.
Секция 11. Эндосимбионты.
Отдел IV . Mendosicutes. Прокариоты, среди которых нет патогенных бактерий; метанобразующие, сероокисляющие, галофилы, микоплазмоподобные, термоацидофильные и другие наиболее древние по происхождению бактерии (архебактерии).
Глава 3. МИР ПРОКАРИОТ
1.
2.
3.
Размеры микроорганизмов
Как показывает само название, объекты, относимые к микроорганизмам, были выделены по признаку их малых размеров. Если принять за критерий границу видимости невооруженным глазом, равную 70–80 мкм (1 миллиметр (мм) = 103 микрометров (мкм) = 106 нанометров (нм), то все объекты, которые лежат за пределами этой границы, можно отнести к микроорганизмам. Мир микроорганизмов – это преимущественно мир одноклеточных форм. Диапазон размеров микроорганизмов велик. Величина самых крупных представителей микромира, лежащих на границе видимости невооруженным глазом, приблизительно 100 мкм (некоторые диатомовые водоросли, спирохеты, высшие протисты). На порядок ниже размеры одноклеточных зеленых водорослей и клеток дрожжей, еще ниже размеры, характерные для большинства бактерий. В среднем линейные размеры бактерий лежат в пределах 0,5–3 мкм, но есть среди бактерий свои «гиганты» и «карлики». Например, клетки нитчатой серобактерии Beggiatoa alba имеют диаметр до 50 мкм; Achromatium oxaliferum , считающийся одним из крупных бактериальных организмов, имеет в длину до 125 мкм при поперечнике примерно 5–33 мкм, а длина клетки спирохеты может быть до 250 мкм.
Самые мелкие из известных прокариотных клеток – микоплазмы, риккетсии и хламидии. Описаны микоплазмы с диаметром клеток 0,1–0,15 мкм. Поскольку молекулы всех соединений имеют определенные физические размеры, то, исходя из объема клетки с диаметром 0,15 мкм, легко подсчитать, что в ней может содержаться порядка 1200 молекул белка и осуществляться около 100 ферментативных реакций. Минимальное число ферментов, нуклеиновых кислот и других макромолекулярных компонентов, необходимых для самовоспроизведения теоретической «минимальной клетки», составляет, по проведенной оценке, около 50. Это то, что необходимо для поддержания клеточной структуры и обеспечения клеточного метаболизма. Таким образом, у группы микоплазм достигнут размер клеток, близкий к теоретическому пределу клеточного уровня организации жизни. Мельчайшие микоплазменные клетки равны или даже меньше частиц другой группы микроскопических организмов – вирусов.
Дата добавления: 2019-09-13; просмотров: 432; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!