МЕХАНІЗМ ВИНИКНЕННЯ ЗБУДЖЕННЯ
Більшість фізіологів виникнення збудження пов'язують з переміщенням іонів і різницею їх концентрації.
Основоположник іонної теорії В. Ю. Чаговець (1896) вважав, що пошкодження або подразнення тканини викликає посилений обмін речовин з утворенням вугільної кислоти, що дисоціюється на іони НСОз і Н+. Іони НСОз, як малорухливі, затримуються в місцях пошкодження, або подразнення, створюючи негативний заряд, а іони Н+, як більш рухливі, переміщуються в сусідні ділянки тканини, утворюючи позитивний заряд. Різниця потенціалів двох сусідніх ділянок тканини і є джерелом електрорушійних сил. Ця теорія отримала назву альтераційної, тобто перемінної (лат. аііегаііо — зміна).
Ю. Беренштейн (1902) висунув мембранну теорію. Він вважав, що поверхня клітинної цитоплазми має напівпроникну мембрану, що в спокійному стані пропускає тільки катіони. В незбудженому стані зовнішня поверхня клітини буде мати позитивний заряд, а внутрішня — негативний. Пошкодження клітини або її подразнення порушують проникність мембрани і вона починає пропускати аніони. Завдяки цьому клітина зовні стає електронегативною. Різниця потенціалів між клітиною та навколишнім середовищем створює умови для виникнення і поширення збудження.
Нерст (1909) причину збудження вбачав у нагромадженні рухливих іонів біля напівпроникних клітинних мембран. При подразненні тканини електричним струмом по обидва боки колоїдної мембрани концентруються певні іони, що здатні викликати збудження.
|
|
|
Ж. Леб (1915) пояснював збудження співвідношенням одно- та двовалентних катіонів. Одновалентні іони калію та натрію підвищують збудливість нервової і м'язової тканин, а двовалентні іони кальцію і магнію, навпаки, знижують її. Таким чином, нагромадження іонів натрію і калію діють подразнювальне, а нагромадження іонів кальцію і магнію — пригнічувально.
П. П. Лазарєв (1916, 1923) на основі досліджень Леба і Нерста про дію іонів на живі клітини дійшов висновку, що одновалентні іони рухаються швидше, ніж двовалентні. Цим зумовлюється підвищення збудливості на катоді та зниження її на аноді у момент замикання гальванічного струму.
За останні роки широкого поширення набула мембранна теорія Ходжкіна—Хакслі (1949), або теорія калієвонатрієвого насосу. Згідно з цією теорією будь-яка жива клітина вкрита мембраною, яка складається з чотирьох шарів молекул. Зовнішній шар — це білкові молекули, далі ідуть два шари фосфоліпідів і четвертий, внутрішній, шар представлений білковими молекулами. Товщина мембрани становить близько 5 А. Один ангстрем дорівнює стомільйонній частині сантиметра (108). У мембрані є “пори” і “канали”, діаметром 7—8 А. Пори несуть певний електричний заряд, який полегшує або ускладнює дифузію іонів через мембрану.
|
|
|
Мембрана більше проникна для катіонів, ніж для аніонів. Різні катіони також проникають через мембрану з різною швидкістю.
За ДОПОМОГОЮ мікроелектродної техніки устаНОВЛЄНО, ЩО електричний потенціал на мембрані позитивний, а всередині
— негативний. Різниця потенціалів пояснюється іонним складом усередині
і зовні клітини (табл.,45). Іони калію порівняно з іонами натрію швидше проникають через мембрану. Академік М. М. Шемякін (1967) вважає, що збільшення проникності мембрани для калію пов'язане з антибіотиком валіноміцином. Вступаючи у сполучення з катіоном калію, валіном'цин утворює позитивно заряджений комплекс, що легко проникає через пори мембрани в середину клітини з негативним електричним зарядом.
Маючи уявлення про структуру та деякі функціональні особливості клітинної мембрани, розглянемо, як виникає і поширюється нервовий імпульс, тобто хвиля збудження.
Первісною реакцією клітини на дію зовнішнього фактора є зміна проникності клітинної мембрани у місці подразнення. Іони натрію миттєво проникають в середину клітини, даючи позитивний заряд. У той же час з клітини виходить незначна кількість іонів калію (рис. 121). За один імпульс клітина втрачає мільйонну частину всієї кількості калію. У результаті іонних переміщень настає деполяризація — клітинний потенціал знижується до нуля. Потім відбувається перезарядка мембрани: усередині плюс, а зобні мінус. У цей час осцилограф реєструє пік потенціалу дії, що перетвориться в слідові потенціали. Пояснюється це повторними змінами проникності мембрани, при якій іони натрію повертаються у міжклітинну рідину, а іони калію — в клітину. Мембрана знову набуває попереднього заряду (зовні позитивний, а всередині негативний), і потенціал спокою відновлюється.
|
|
|
Переміщення іонів натрію в середину клітини є причиною появи місцевого кругового струму, який поширюється на поверхні мембрани. В середині клітини круговий струм іде від збудженої ділянки до незбудженої, а зовні — від місця спокою до збудженого. Проникаючи в сусідню незбуджену ділянку, місцевий струм відкриває пори для натрію, створюючи тут новий імпульс. Зароджуючись у кожній наступній точці нервового або м'язового волокна, імпульс поширюється на значну відстань.
|
|
|
Отже, нервовий імпульс — складний електрохімічний процес. Що ж є причиною активного переміщення через мембрану іонів калію і натрію? У дослідах на аксоні кальмара, товщина якого досягає 1 мм, доведено, що необхідна енергія для “іонного насосу” утворюється в процесі “спалювання” аденозинтрифосфорної кислоти (АТФ).
ФАЗИ ЗБУДЛИВОСТІ
У процесі виникнення й розвитку одинокого імпульсу збудження мають місце фазові зміни збудливості тканини. Спочатку після нанесення подразнення спостерігається короткотривале, незначне місцеве підвищення збудливості. Потім настає різке зниження збудливості. Ця тимчасова незбудливість отримала назву абсолютної рефрактерної фази. Вона відповідає розвитку потенціалу дії.
Тривалість фази абсолютної рефрактерності дорівнює:
для нерва — 0,002 с, скелетного м'яза — 0,005 і серцевого м'яза — 0,3—0,4 с.
Після закінчення періоду абсолютної рефрактерності збудливість тканини поступово відновлюється, досягаючи початкового рівня. Цей період зниженої збудливості називається ф а з о ю, відносної незбудливості (рефрактерності). Ця фаза в часі збігається з переходом піку потенціалу у слідову електронегативність. У фазі відносної рефрактерності подразники великої сили можуть викликати відповідь. Тривалість її для нервових волокон становить 0,001—0,01, а для м'язів — 0,03 с.
Далі настає фаза підвищеної збудливості, яку М. Є. Введенський назвав екзальтаційною. Ця фаза збігається з слідовою електронегативністю. У нерві вона триває 0,02, а у м'язі 0,05 с і більше.
Після фази екзальтації настає тривала фаза субнор-мальності, протягом якої збудливість тканини дещо знижується порівняно з початковою збудливістю. За часом дана фаза збігається з електропозитивністю. Потім збудливість тканини приходить до норми.
З вищесказаного виходить, що всі фази зміни збудливості відповідають певним періодам потенціалу дії (рис. 122).
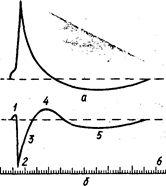
Знання фазових змін збудливості важливе для розуміння характеру скорочення м'язів залежно від частоти подразнення. Якщо подразнювати нерв нервово-м'язового препарату індукційним струмом за умови, що друге подразнення іде за першим з інтервалом, рівним тривалості абсолютної рефрактерності, м'яз не скоротиться. Поступове збільшення інтервалу буде супроводжуватись збільшенням висоти скорочення. Максимальне скорочення м'яза настає в той момент, коли подразнення збігається у часі з екзальтаційною фазою. Тривалість фазових змін збудливості у процесі життєдіяльності тканин змінюється. Так, при втомі м'яза час абсолютної і відносної рефрактерності збільшується. Фази збудження тісно пов'язані з обміном речовин.
Незначне підвищення збудливості на початку збудження стимулює перехід тканини від видимого спокою до діяльного активного стану. У період абсолютної рефрактерності, вперше виявленої на серці, утворюється необхідна енергія, яка забезпечує розвиток збудження. У цей час збільшується проникність мембрани клітини для іонів калію і припиняється потік іонів натрію в клітину. Згідно з мембранною теорією збудження Ходжкіна—Хакслі ці процеси гальмують виникнення нового потенціалу дії на чергове подразнення. Біологічне значення, фази абсолютної рефрактерності полягає в тому, що вона запобігає надмірним втратам енергетичних матеріалів у тканині і тим самим сприяє нормальному розвитку процесу збудження. Відносна рефрактерність створює умови для відновлення вихідного стану збудженої тканини.
Під час фази екзальтації тканина здатна відповідати на повторне подразнення. Поява цієї фази свідчить про те, що подразнення викликає не тільки певну реакцію тканини, а й створює сприятливі умови для повторних збуджень. Фаза екзальтації має безпосереднє відношення до ритмічної діяльності, властивої нервовій та м'язовій тканинам.
Дата добавления: 2019-02-12; просмотров: 103; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!