Тема: Программируемая клеточная смерть.
1. В чем заключается значение апоптоза как общебиологического явления?
2. Назовите функциональные группы молекул, участвующих в процессе апоптоза и охарактеризуйте их.
3. Какие факторы вызывают изменения митохондрии, приводящие к гибели клетки?
4. Как белок р-53 регулирует клеточный цикл?
5. Дайте определение понятий онкогенный вирус, онкоген, протоонкоген, антионкоген (или ген-супрессор опухолей, ГСО) и раскройте молекулярный механизм канцерогенеза.
6. Заполните пропуски в следующих утверждениях.
A. Клетки злокачественных опухолей проникают в другие ткани тела, давая начало вторичным опухолям, называемым _________.
Б. Причиной возникновения опухолей некоторых типов служат _________, инфицирующие нормальные клетки.
B. Клетки, не испытывающие социального контроля деления и вытесняющие благодаря более быстрому росту нормальные клетки, называют _________.
Г. Вирус саркомы Рауса относится к классу ___________.
Д. Некоторые гены, называемые ___________, при попадании в нормальные клетки могут трансформировать их в раковые клетки.
Е. Трансформирующий ген вируса саркомы Рауса обозначается _________.
Ж. Гены нормальных клеток, гомологичные онкогенам, называют ________.
7. Укажите, какие из следующих утверждений правильные, а какие – нет. Если утверждение неверно, объясните почему.
А. Для трансформированных клеток характерен целый ряд признаков, которые можно разбить на три группы: аномалии, связанные с плазматической мембраной, аномалии прикрепления и аномалии роста и деления.
|
|
|
Б. Трансформация клетки может быть вызвана одним геном.
В. Температурочувствительные мутанты по гену v- src из вируса саркомы Рауса были использованы для демонстрации того, что трансформирующий ген действует как переключатель в определенной точке клеточного цикла и в другое время не нужен для поддержания трансформированного состояния.
Г. При анализе трансформированных клеток на онкогены выявляются только доминантные онкогены.
Д. Рецессивные мутации генов социального контроля могут быть более распространенной причиной рака, чем доминантные мутации.
Е. В сущности все вирусные онкогены соответствуют либо естественным факторам роста, либо рецепторам естественных факторов роста.
Ж. По-видимому, большую роль в трансформации клеток играет тирозинспецифическая протеинкиназная активность, которой обладает продукт гена v- src вируса саркомы Рауса.
З. Продукт гена v- src прикрепляется к плазматической мембране коротким отрезком из гидрофобных аминокислот на N-конце полипептидной цепи.
И. Каким образом пролиферация клеток связана с их адгезией, неизвестно.
|
|
|
К. Нормальные клетки делятся только тогда, когда они плотно прикреплены к субстрату, а пролиферация трансформированных клеток существенно тормозится, если искусственно вызвать их прикрепление к субстрату.
Л. Одинаковые размеры органов и тела у тритонов с разной плоидностью указывают на то, что необходимый для регуляции размеров тела контроль клеточного деления может зависеть от «измерения» длины, а не от простого «подсчета» числа клеток или циклов деления.
8. Было высказано предположение, что ограниченность числа делений нормальных клеток (до примерно 50 циклов) лимитирует максимальные размеры опухолей и тем самым обеспечивается некоторая защита от рака. Приняв, что 108 клеток имеют массу 1 г, рассчитайте массу опухоли, возникающей при 50 делениях одной раковой клетки.
9. Ретинобластома – это чрезвычайно редкая разновидность рака нервных клеток глаза. Заболевание поражает преимущественно детей в возрасте до пяти лет, поскольку оно может возникнуть только тогда, когда нервные клетки еще делятся. Иногда опухоль поражает лишь один глаз, но бывает и так, что опухоли развиваются на обоих глазах. В последнем случае болезнь всегда носит наследуемый характер, тогда как поражение одного глаза чаще всего встречается в семьях, где раньше этого заболевания не было.
|
|
|
Если построить график зависимости той части детей, у которых заболевание ещё не возникло, от возраста детей, то выявляется весьма информативное различие между двухсторонним и односторонним поражениями (рис. 11-1). В первом случае плавный ход кривой свидетельствует о том, что для возникновения двухсторонней ретинобластомы достаточно одного события. Напротив, наличие «плеча» на кривой для односторонней ретинобластомы говорит о том, что для возникновения такого поражения в одном нейроне должно произойти несколько событий. (Плечо указывает на то, что события накапливаются постепенно. Например, если необходимы два события, то в большинстве клеток в ранний период времени происходит только одно событие, что не приводит к возникновению опухоли. С течением времени возрастает вероятность, что в уже пораженной клетке случится второе событие и, следовательно, возникнет опухоль.)
Для объяснения этих наблюдений можно предположить следующее. Опухоль развивается в том случае, если обе копии гена ретинобластомы ( Rb ) утрачены или содержат мутации. В случае наследственной (двухсторонней) формы этого заболевания ребенок получает дефектный ген Rb от одного из родителей, и если в какой-либо нервной клетке одного глаза другая копия гена утрачивается в результате соматической мутации, то в этом глазу возникает опухоль. В действительности утрата копии гена случается довольно часто, поэтому опухоли обычно развиваются в обоих глазах. Если ребенок получает две хорошие копии гена Rb , то опухоль в глазу возникнет только в том случае, если в одной и той же клетке будут утрачены обе копии. Поскольку такая двойная потеря случается очень редко, то опухоль появляется обычно только на одном глазу.
|
|
|
|
|

Чтобы проверить эту гипотезу, вы исследуете с помощью клона кДНК гена Rb структуру гена в клетках от здоровых людей и от больных с односторонней и двухсторонней ретинобластомой. Как показано на рис. 11-2, у здоровых людей имеются четыре рестрикционных фрагмента, которые гибридизуются с кДНК-зондом (это означает, что каждый из этих фрагментов содержит по крайней мере один экзон). Фибробласты (неопухолевые клетки) от обоих больных также содержат эти четыре фрагмента, хотя количество трех фрагментов от ребенка с двухсторонней ретинобластомой вдвое меньше, чем в норме. В опухолевых клетках обоих больных отсутствуют некоторые рестрикционные фрагменты.
Рис. 11-1. Время появления признаков односторонней и двухсторонней ретинобластомы. На графике представлены данные для популяции детей, у которых в том или ином возрасте развивается ретинобластома. Показана зависимость той доли популяции, в которой ретинобластома еще не возникла, от времени после рождения.
|
|
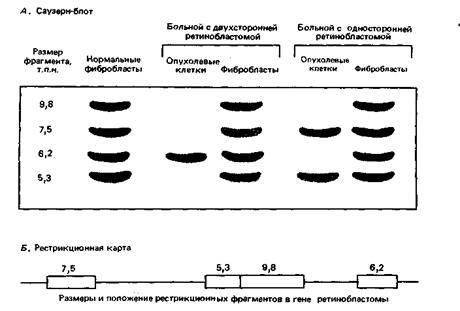
Рис. 11-2. Результаты блот-гибризации рестрикционных фрагментов гена ретинобластомы.
А. Картины гибридизации (Саузерн-блот) у здоровых людей в случаях заболевания односторонней или двухсторонней ретинобластомой. Более светлая окраска полос указывает на вполовину меньшее, чем в норме, число копий. Б. Расположение рестрикционных фрагментов. Фрагменты, держащие экзоны (показанные в виде прямоугольников), гибридизуются с кДНК, использованной в качестве зонда в этих опытах.
A. Объясните, почему фибробласты и опухолевые клетки от одного и того же больного дают разные распределения полос?
Б. Какова структура генов Rb в фибробластах и опухолевых клетках от двух больных?
B. Согласуются ли эти результаты с гипотезой о том, что ретинобластома обусловлена утратой гена Rb?
Г. Каким образом отсутствие продукта гена Rb может быть причиной развития ретинобластомы?
11. Цитокинез – процесс собственно клеточного деления – привлекает внимание ученых на протяжении более чем ста лет. Предложены всевозможные объяснения цитокинеза; задача состоит в том, чтобы решить, какое из них является верным. Рассмотрим следующие три гипотезы цитокинеза.
1. Хромосомная сигнализация. Хромосомы при расщеплении в анафазе испускают сигнал, воспринимаемый близлежащей клеточной поверхностью и инициирующий образование борозды деления.
2. Полюсная релаксация. Звезды снимают натяжение в ближайших к ним участках клеточной поверхности (области полюсов), что дает возможность участку мембраны, наиболее удаленному от полюсов (в экваториальной плоскости), сократиться и начать образование борозды деления.
3. Стимуляция звездами. Звезды стимулируют сокращение в районе клеточной поверхности, где перекрываются противоположно направленные нити веретена (т. е. в экваториальной плоскости), вызывая таким образом образование борозды деления.
Эти гипотезы проверяли многими способами. Один особенно информативный эксперимент состоял в том, что стеклянный шарик вдавливали в центр дробящегося яйца плоского морского ежа так, чтобы оно деформировалось с образованием тора (форма бублика). При первом делении, как показано на рис. 11-3, образуется одна колбасовидная клетка, при втором делении она разделяется на четыре клетки.
|
|

Рис. 11-3. Первое и второе деления тороидального яйца плоского морского ежа.
Занятие 12
Тема: Генетическая инженерия
1. Заполните пропуски в следующих утверждениях.
А. ____________ разрезают двухцепочечную спираль ДНК по специфическим последовательностям, состоящим из четырех-восьми нуклеотидов (палиндромы), разделяя ее на фрагменты строго определенных размеров, которые называются ________.
Б. С целью размножения (амплифицирования) и получения в чистом виде тех или иных генов фрагменты эукариотической ДНК могут быть встроены в специально подготовленные бактериальные вирусы или в плазмиды, называемые _________.
В. Колония клеток, происходящая от одной клетки, называется ____________ клеток.
Г. Отдельная колония бактерий, которые содержат плазмиду с включенным в нее фрагментом ДНК человека, называется ___________; набор таких бактерий, представляющий весь геном человека, составит ___________.
Д. Двухцепочечные ДНК-копии эукариотических мРНК, полученные с помощью обратной транскриптазы, могут быть встроены в векторы клонирования для создания _________, которая может использоваться для выявления отдельных клонов, соответствующих определенным мРНК.
Е. Если от одного организма имеются клетки двух близких между собой типов и при этом интересующий исследователя белок или белки производят клетки только одного типа, то перед клонированием кДНК можно прибегнуть к методу __________, позволяющему обогатить смесь определенными нуклеотидными последовательностями.
Ж. Использование перекрывающихся клонов для продвижения от известного гена к ближайшему гену называется методом _________.
З. Чтобы установить, содержит ли данный клон ДНК, кодирующую ранее охарактеризованный белок, часто проводят _________, т.е. клонированную ДНК используют для того, чтобы отобрать соответствующую мРНК, продукт трансляции которой можно затем сравнить с известным белком.
И. Плазмиды или вирусы с сильными промоторами, размещенными так, что продукт клонируемого гена может производиться в большом количестве, называются ____________ векторами.
К. Новый гибридный ген, который кодирует __________, может быть образован путем соединения частей кодирующих последовательностей двух разных генов.
Л. В клетках зародышевой линии у ________ мышей присутствуют хромосомы со стабильными изменениями, которые произошли в результате интеграции клонированной ДНК.
М. Очищенные ДНК-полимеразы и синтетические олигонуклеотиды ДНК можно использовать для амплификации отдельных областей генома с помощью метода ________.
2. Укажите, какие из следующих утверждений правильные, а какие – нет. Если утверждение неверно, объясните почему.
А. Не все рестрицирующие нуклеазы производят ступенчатые разрывы в ДНК.
Б. Плазмидными векторами, используемыми при клонировании, могут быть небольшие молекулы, которые содержат уникальные сайты рестрикции, чтобы включать чужеродную ДНК, имеют свою точку начала репликации ДНК, а также ген, сообщающий клетке устойчивость к какому-либо антибиотику.
В. Библиотеки кДНК содержат только те последовательности, которые экспрессировались в ткани, откуда была выделена исходная мРНК, тогда как библиотеки геномных ДНК содержат набор всех последовательностей, имеющихся в организме.
Г. Аминокислотную последовательность эукариотического белка можно одинаково хорошо определить как при помощи клона кДНК, так и при помощи геномного клона.
Д. Для очистки особых типов редких мРНК можно применить метод иммунопреципитации полисом, используя специфические антитела против рибосом.
Е. Субтрактивная гибридизация представляет собой способ для обогащения теми мРНК, которые присутствуют в клетках одного типа и отсутствуют в клетках другого типа у одного и того же организма.
Ж. Чтобы получить большое количество эукариотического белка в клетках бактерий, в экспрессирующий вектор с сильным промотором помещают геномный клон, содержащий целый ген.
З. Трансгенных животных можно создавать путем микроинъекции ДНК в ядро соматической клетки, где ДНК может включаться в хромосому и таким образом сохраняться и экспрессироваться в последующих клеточных поколениях.
3. Рестриктазы ВаmНI и РstI разрезают узнаваемые ими последовательности, как показано на рис. 12-1.
А. Укажите, где находятся 5'- и 3'-концы разрезаемых молекул ДНК.
Б. Как будут модифицироваться эти концы, если инкубировать разрезанные молекулы ДНК с ДНК-полимеразой в присутствии всех четырех дезоксинуклеозидтрифосфатов?
В. Могут ли концы, появившиеся в результате разрезания ферментом ВаmНI, соединиться вновь при инкубации с ДНК-лигазой фага Т4 после того, как была проведена реакция, о которой шла речь в пункте Б?
Можно ли соединить вместе концы ДНК, образовавшиеся при обработке РstI? (Лигаза ДНК фага Т4 соединяет вместе «тупые» концы так же, как и «липкие».)
Г. Будет ли регенерировать узнаваемый ВаmНI сайт, о котором шла речь в пункте В, при соединении концов? Будет ли регенерировать сайт, узнаваемый РstI?
|
|

Рис. 12-1. Расщепление рестриктазами ВаmНI и РstI узнаваемых ими последовательностей. Показаны только те нуклеотиды,которые образуют сайты узнавания.
4. Вы очистили два фрагмента ДНК, образующиеся при обработке ВаmНI рекомбинантной плазмидной ДНК. Один фрагмент содержит 400 п. н., другой – 900 п. н. Вы хотите объединить их вместе, как показано на рис. 12-2, чтобы создать гибридный ген, который в том случае, если ваши ожидания правильны, будет иметь новые удивительные свойства.
Вы смешиваете два фрагмента в присутствии ДНК-лигазы и инкубируете смесь. Через 30 мин после начала инкубации и второй раз через 8 ч. вы отбираете пробы и анализируете их методом электрофореза в геле. Вас удивляет, что вместо ожидаемой рекомбинантной молекулы размером 1,3 т. п. н. электрофореграмма содержит сложный набор фрагментов (рис. 12-3, А). Вы замечаете, что при более длительной инкубации интенсивность полос, соответствующих более мелким фрагментам, уменьшается, а интенсивность полос, соответствующих более крупным фрагментам, увеличивается. Если вы добавляете к смеси сшиваемых молекул ВаmНI, то вновь образуются исходные фрагменты (рис. 12-3, А).
Находясь в недоумении, но настойчиво продолжая исследование, вы проводите очистку фрагмента размером 1,3 т. п. н. с помощью гель-электрофореза, контролируя структуру путем обработки пробы рестриктазой ВаmНI. Как вы и ожидали, при этом заново появляются две исходные полосы (рис. 12-3, Б). Чтобы убедиться все-таки, что это именно та структура, которую вы хотели получить, вы обрабатываете другую пробу рестриктазой ЕсоRI. Вы ожидали, что при этом получатся два фрагмента длиной 300 нуклеотидов и один фрагмент размером 700 нуклеотидов. И снова вы удивлены сложностью набора полос в геле (рис. 12-3, Б).
|
|

Рис. 12-2. Окончательная структура гибридного гена.
|
|

Рис. 12-3. Сшивание фрагментов очищенной ДНК (А) и диагностическое расщепление очищенного фрагмента длиной 1,3 т. п. н. (Б).
А. Почему исходная смесь соединяющихся фрагментов ДНК дает так много полос в геле?
Б. Почему при обработке очищенного фрагмента длиной 1,3 т. п. н. рестриктазой ЕсоRI образуется так много фрагментов?
5. Вы провели клонирование сегмента (4000 п. н.) интересующего вас гена в плазмидном векторе (рис. 12-4) и теперь хотите получить карту рестрикции этого гена, чтобы перейти затем к другим манипуляциям с ДНК. Ваш руководитель оставил вам инструкции относительно того, как все это нужно сделать, и отправился в отпуск, так что вы можете полагаться только на себя. Вы следуете его инструкциям, перечисленным ниже.
1. Разрезать плазмиду рестриктазой ЕсоRI.
2. Добавить к концам, образовавшимся после обработки ЕсоRI, радиоактивную метку.
3. Разрезать меченую ДНК рестриктазой ВаmНI.
4. Очистить встроенную ДНК от плазмиды.
5. Обработать меченую ДНК-вставку рестриктазой в течение короткого периода времени так, чтобы каждая меченая молекула была разрезана в среднем один раз.
6. Повторить этап 5, используя разные рестриктазы.
7. Провести электрофорез частично расщепленных ДНК в агарозном геле, причем препараты, обработанные разными рестриктазами, нанести на гель рядом друг с другом.
8. Наложить гель на рентгеновскую пленку, чтобы получился радиоавтограф фрагментов с радиоактивным концом.
9. Нарисовать рестрикционную карту.
Наиболее трудным для вас был этап 5, однако, уменьшая количество фермента и снижая температуру, вам удалось подобрать условия для частичного расщепления. Затем вы смогли полностью выполнить этап 8 и получить радиоавтограмму, представленную на рис. 12-5.
К сожалению, руководитель не объяснил вам достаточно подробно, как строить карту по данным радиоавтограммы. Он должен вернуться завтра. Хватит ли вам времени, чтобы нарисовать ее?
|
|
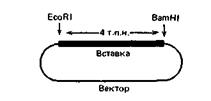
Рис. 12-4. Рекомбинантная плазмида, содержащая клонированный сегмент ДНК.
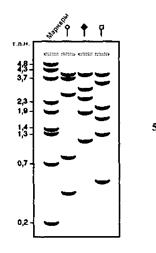
Рис. 12-5. Радиоавтограмма, показывающая электрофоретическое разделение меченых фрагментов после частичного расщепления тремя рестриктазами, обозначенными разными символами. Числа слева указывают размеры маркерных фрагментов (в т. п.н.).
6. Полночь. Вас разбудил коллега, чтобы поделиться еще одним грандиозным проектом. Он потратил два последних года на очистку белка, представляющего собой сильный модулятор иммунного ответа. Вечером он получил первые 30 аминокислот на аминокислотном анализаторе (рис. 12-6). Ему нужен ваш совет, как лучше клонировать ген, чтобы добиться высокого уровня его экспрессии в бактериях. Он доказывает, что этот белок, благодаря стимулированию имунной системы, мог бы служить отличным средством для лечения простуды. Он и название ему уже подобрал – иммустим.
Хотя коллега и увлекающийся человек, но он ваш друг, поэтому вы откликаетесь на его идею, пообещав позвонить через 15 мин, как только переведете аминокислотную последовательность в нуклеотидную. Какие два набора олигонуклеотидов размером по 20 нуклеотидов вы порекомендуете другу-коллеге в качестве наилучших зондов гибридизации для скрининга библиотеки геномной ДНК?
|
|

Рис. 12-6. Первые 30 аминокислот, определенные вашим коллегой с помощью аминокислотного анализатора
7. На основе своей предыдущей работы вы предполагаете, что глутамин (Q) в сегменте белка, изображенном на рис. 12-7, играет важную роль в активном центре. Ваш руководитель хочет, чтобы вы изменили белок тремя путями: заменили глутамин на лизин (К), заменили глутамин на глицин (G) и просто удалили глутамин из белка. Вы планируете произвести эти мутационные изменения путем гибридизации соответствующего олигонуклеотида с ДНК вируса М13. Когда олигонуклеотид расположится вдоль кольцевой молекулы М13, с участием ДНК-полимеразы будет достроена полная цепь, кодирующая комплемент желаемого мутантного белка. В качестве первого шага в осуществлении мутационных изменений предложите три олигонуклеотида размером 20 нуклеотидов, которые могли бы гибридизоваться с клонируемым геном на одноцепочечной ДНК вируса М13.
|
|
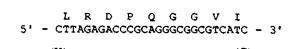
Рис. 12-7. Последовательности ДНК и кодируемого ею белка
Дата добавления: 2018-10-25; просмотров: 1448; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!