Схема шляхів обміну безнітрогенових залишків амінокислот в організмі, зв'язок з циклом Кребса.
В ходе катаболизма аминокислот происходит отщепление аминогруппы и выделение аммиака. Другим продуктом дезаминирования аминокислот служит их безазотистый остаток в виде α-кетокислот. Катаболизм аминокислот происходит практически постоянно. За сутки в норме в организме человека распадается примерно 100 г аминокислот, и такое же количество должно поступать в составе белков пищи.
Большая часть безазотистых остатков аминокислот превращается в пируват либо непосредственно (Ала, Сер), либо в результате более сложного пути, превращаясь вначале в один из метаболитов ЦТК. Затем в реакциях цитратного цикла происходит образование оксалоацетата, который превращается в фосфоенолпируват. Из фосфоенолпирувата под действием пируваткиназы образуется пируват. Пируват подвергается окислительному декарбоксилированию и превращается в ацетил-КоА, который окисляется в ЦТК до СО2 и Н2О с выделением энергии. Такой путь проходят преимущественно аминокислоты пищи.
При недостатке глюкозы в организме фосфоенолпируват включается в глюконеогенез (см. раздел 7). Это происходит при голодании, длительной физической работу при сахарном диабете и других тяжёлых хронических заболеваниях, сопровождающихся распадом собственных белков организма. Скорость глюконеогенеза из аминокислот регулируется гормонами. Так, под действием глюкагона увеличивается активность регуляторных ферментов процесса, а кортизол индуцирует синтез ферментов глюконеогенеза в печени. Активация глюконеогенеза из аминокислот происходит и при преимущественно белковом питании.
Глікогенні та кетогенні амінокислоти.
Катаболизм всех аминокислот сводится к образованию шести веществ, вступающих в общий путь катаболизма: пируват, ацетил-КоА, α-кетоглутарат, сукцинил-КоА, фумарат, оксалоацетат
Аминокислоты, которые превращаются в пируват и промежуточные продукты ЦТК (α-КГ, сукцинил-КоА, фумарат) и образуют в конечном итоге оксалоацетат, могут использоваться в процессе глюконеогенеза. Такие аминокислоты относят к группе гликогенных аминокислот.
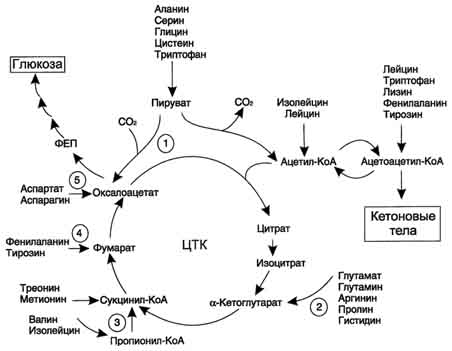
Некоторые аминокислоты в процессе катаболизма превращаются в ацетоацетат (Лиз, Лей) или ацетил-КоА (Лей) и могут использоваться в синтезе кетоновых тел. Такие аминокислоты называюткетогенными.
Ряд аминокислот используется и для синтеза глюкозы, и для синтеза кетоновых тел, так как в процессе их катаболизма образуются 2 продукта - определённый метаболит цитратного цикла и ацетоацетат (Три, Фен, Тир) или ацетил-КоА (Иле). Такие аминокислоты называют смешанными, илигликокетогенными(рис. 9-22, табл. 9-5).
Таблица 9-5. Классификация аминокислот по судьбе безазотистого остатка
| Гликогенные аминокислоты | Гликокетогенные аминокислоты | Кетогенные аминокислоты |
| Алании | Тирозин | Лейцин |
| Аспарагин | Изолейцин | Лизин |
| Аспартат | Фенилаланин | |
| Глицин | Триптофан | |
| Глутамат | ||
| Глутамин | ||
| Пролин | ||
| Серии | ||
| Цистеин | ||
| Аргинин | ||
| Гистидин | ||
| Валин | ||
| Метионин | ||
| Треонин |
Обмін фенілаланіну та тирозину, порушення, шляхи корекції.
Фенилаланин относится к незаменимым аминокислотам, поскольку ткани животных не обладают способностью синтезировать его бензольное кольцо. В то же время тирозин полностью заменим при достаточном поступлении фенилаланина с пищей. Объясняется это тем, что основной путь превращенияфенилаланина начинается с его окисления (точнее, гидрокси-лирования) в тирозин (рис. 12.6). Реакциягидроксилирования катализируется специфической фенилаланин-4-монооксигеназой, которая в качествекофермента содержит, как все другие гидроксилазы, тетрагидро-биоптерин. Блокирование этой реакции, наблюдаемое при нарушении синтеза фенилаланин-4-монооксигеназы в печени, приводит к развитию тяжелой наследственной болезни – фенилкетонурии (фенилпировиноградная олигофрения). В процессетрансаминирования тирозин превращается в n-оксифенилпировиноградную кислоту, которая под действием специфической оксидазы подвергается окислению, декарбоксилированию, гидро-ксилированию и внутримолекулярному перемещению боковой цепи с образованием гомогентизиновой кислоты; эта реакциятребует присутствия аскорбиновой кислоты, роль которой пока не выяснена. Дальнейшее превращениегомогентизиновой кислоты в малеилацетоуксусную кислоту катализируется оксидазой гомогентизиновой кислоты. Малеилацетоуксус-ная кислота под действием специфической изомеразы в присутствии глу-татиона превращается в фумарилацетоуксусную кислоту, подвергающуюся гидролизу с образованием фумаровой и ацетоуксусной кислот, дальнейшие превращения которых уже известны.
Дата добавления: 2018-06-01; просмотров: 421; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!