Потенціалом дії (ПД) називають швидке коливання МПС, що виникає під час збудження клітини.
ЛЕКЦІЯ 2
ФІЗІОЛОГІЯ ЗБУДЛИВИХ ТКАНИН
План
1. Основні властивості збудливих тканин ср
1.1. Подразники. Класифікація подразників ср
1.2. Показники виміру збуджуваності
1.3. Теорія Ходжкіна -Хакслі
2. Мембранний потенціал спокою
3. Мембранний потенціал дії
3.1. Зміни збудливості клітини під час збудження
3.2. Іонні канали
4. Біоструми ср
4.1. Лабільність
4.2. Ритми подразнення
4.3. Парабіоз
5. Фізіологія нервів
5.1. Будова нервової системи. М’якотні та безм’ְякотні нервові волокна ср
5.2. Властивості нервових волокон
6. Фізіологія м'язів
6.1. Гістологічна будова скелетних м’язів ср
6.2. Основні властивості скелетних м’язів
6.3. Теорія м’язового скорочення
6.4. Робота м’язів
7. Синапси
Основні властивості збудливих тканин
Основною властивістю живих систем є здатність відповідати на вплив навколишнього середовища активною реакцією. Особливо яскраво ця властивість виявляється в реакціях нервової, м'язової і залозистої тканин, які називають збудливими, тобто здатними відповідати збудженням на вплив подразників. Подразнення зумовлює в клітині складний комплекс мікроструктурних перебудов, а також зміни обміну речовин, концентрації йонів, швидкості їх руху та їх розподілу на клітинних мембранах.
Збудливість - властивість тільки збудливих тканин реагувати на подразнення процесом збудження, поширення якого по збудливих тканинах призводить до прояву специфічної реакції. Збудливість залежить від ряду факторів:
|
|
|
1) приналежності тканини ( нервова - м'язова - залозиста );
2) у межах однієї тканини - від її положення, наприклад залозиста тканина - слинні, підшлункова - шлункові і кишкові залози;
3) від ступеня диференційованості тканини (м'язова тканина литкового м'яза людини більш збудлива, чим литковий м'яз жаби );
4) від температури тіла;
5) від фізіологічного стану організму;
6) від застосування хімічних речовин - малі дози підвищують збудливість, великі - знижують.
Збудливі тканини на подразнення відповідають процесом збудження.
Збудження - складний біологічний процес, при якому в тканинах відбувається ряд змін. Олексій Олексійович Ухтомський ( 1875 - 1942 ) визначав збудження, як перехід тканини від видимого спокою до діяльного стана і самий діяльний стан який продовжується до тих пір, поки тканина знову не повернеться до спокою.
Більш розширене визначення збудження таке:
Збудження - складний біологічний процес, що виникає в збудливих тканинах під впливом дії подразника. Супроводжується зміною обміну речовин, фізико-хімічних властивостей тканин, появою біотоків і інших змін, що призводять до відповідної реакції, специфічної лише для даного виду тканини.
|
|
|
Збудження виникає тільки в таких умовах:
1. При визначеній силі подразника.
2. При визначеній швидкості нанесення подразнення.
3. При визначеній тривалості дії подразника.
Що ж таке подразник?
Подразником називається фактор зовнішнього або внутрішнього середовища, спроможний викликати відповідну реакцію живого організму, або його тканин і клітин. Подразники можна класифікувати за різноманітними ознаками:
1. За місцем дії - зовнішні, внутрішні (нервові імпульси, гормони, продукти обміну речовин ).
2. За біологічною дією на тканину - адекватні, неадекватні.
3. За природою подразників (це, в основному, стосується зовнішніх подразників) механічні, термічні, хімічні, біологічні і електричні.
Найбільш широке поширення одержав в умовах експерименту електричний подразник, що легко дозується, швидко включається і виключається і діє на тканини, не викликаючи в них видимих змін.
За силою подразники діляться:
1) підпорогові;
2) порогові;
3) надпорогові;
3.1.) оптимальні;
3.2.) надмаксимальні (позамежні, песимальні).
До підпорогових подразників відносяться подразники такої сили, що не спроможні викликати відповідну реакцію.
|
|
|
Всі надпорогові подразники завжди викликають відповідну реакцію, проте, відповідна реакція буде різною, у залежності від сили подразника - оптимальні за силою подразники викликають максимальне збудження, надмаксимальні - песимальну відповідь.
Крім визначеної сили подразника для розвитку збудження важлива і швидкість нанесення подразнення або, краще сказати, градієнт подразнення. Градіент подразнення - це швидкість зростання сили подразника.
При повільному збільшенні сили дії подразника збудження може і не виникнути. Так, при повільному здавлювані нерва відбувається подразнення його, але збудження не виникає. При повільному нагріванні м'яза або нерва, тканина може зваритися, не переходячи в стан збудження. Пристосування збудливої тканини до повільно наростаючої сили подразника називають акомодацією. Різноманітні тканини, клітини і їхні структурні елементи (рецептори ) мають різноманітну акомодацію. При максимально швидкому наростанні сили подразника, поріг збудження знижується. Тривалість дії подразника має визначену залежність від сили подразника.
Показники виміру збудливості
Для виміру збуджуваності можна користуватися декількома показниками, один із них - гранична сила подразника. Показник граничної сили дії подразника характеризує фізіологічний стан збудливої тканини. Чим збуджуваність тканини вище, тим нижче гранична сила для розвитку процесу збудження, і навпаки. Мінімальну силу подразника, спроможну викликати збудження, називають реобазою. Якщо побудувати графік, у якому на осі ординат буде відкладена сила подразника, а на осі абсцис - мінімальний час її дії, тоді одержимо криву сили часу. Найменший період часу, протягом якого діє струм силою в одну реобазу, щоб викликати потенціал дії, називається корисним часом.
|
|
|
Є визначена залежність між силою і тривалістю дії подразника. Чим сильніше подразник, тим коротше час його дії, необхідний для виникнення збудження. Проте при силі подразника в одну реобазу, навіть на дуже чутливих приладах, важко визначити корисний час, тому що незначним змінам сили буде відповідати великі зміни в часу.
Піддавши математичному аналізу криву сили – тривалості, Л. Лапик приходив до висновку, що для характеристики збудливості за часом дії подразника потрібно взяти час дії подвоєної граничної сили цього подразника. Тоді точка, що відповідає часу дії подвоєної граничної сили, буде знаходитися в місці перегину (крутого вигину кривої; у цьому випадку тривалість подразнення визначається точно). Отже, більш точно збуджуваність буде характеризуватися показником, називаним хронаксією.
ХРОНАКСІЯ - мінімальний час, необхідний для виникнення збудження тканини при дії подвоєної реобази (подвоєної граничної сили подразника). Хронаксія виражається в тисячах частках секунди (у мілісекундах), або сигмах. Чим менше хронаксія, тим вище збуджуваність тканини. Хронаксію тканин вимірюють спеціальним приладом – хронаксометром. Спочатку визначають реобазу, потім її подвоюють і визначають мінімальний час дії подвоєної реобази, необхідне для виникнення збудження. Вимір хронаксії проводять не тільки в експериментальній фізіології, але й у клініці для об'єктивної оцінки фізіологічного стану тканини або органа.
0-а – хронаксія
0-в – корисний час
0-1 – реобаза
0-2 подвійна реобаза
Корисний час – найменший період, протягом якого діє струм силою в одну реобазу, щоб викликати потенціал дії.
Дослідження показали, що хронаксія є перемінною. Розмір її залежить від структури тканини, від стана тканини, органа і всього організму в цілому. Хронаксія рухових нервів менше, ніж скелетних м'язів. Наприклад, у коня і жуйних хронаксія рухових нервів коливається від 0,09 до 0,2 мілісекунди, хронаксія скелетних м'язів - від 0,2 до 0,4 мілісекунди, при чому хронаксія згиначів у 1,5 - 2 рази менше, чим розгиначів. Нерви вегетативної нервової системи мають дуже високу хронаксію (біля 5 мілісекунд). Найвища хронаксія, що вимірюється не мілісекундами, а сотими і десятими частками секунди, у гладких м'язів шлунка, кишечника і матки.
Якщо при подразненні процес розвивається тільки в місці нанесення подразнення і не поширюється по тканині, то збудження - процес, що поширюється, виниклий в однім місці він поширюється по всій тканині або органу, а іноді переходить і на сусідні тканини й органи. Як здійснюється перехід із тканини на тканину ми розберемо пізніше, а зараз розберемо, що ж являє собою процес збудження і як він здійснюється.
Збудливих тканин в організмі тварин нараховується три види. Це нервова, м'язова і залозиста тканина. Процеси, що відбуваються в них, при збудженні в принципі не відрізняються, хоча і мають розбіжності в деталях. Як і всі інші тканини, вони оточені клітинною мембраною, що має визначений мембранний потенціал, причому на відміну від незбудливих тканин потенціал мембрани досить високий (табл.).
| Вид тканини | Мембранний потенціал, мВ |
| Епітеліальні клітини | 15 - 35 |
| Клітини опорно-трофічної тканини | 30 - 50 |
| Нервові клітини і волокно | 60 - 70 |
| Поперечносмугасте м'язове волокно | 60 - 90 |
Теорія Ходжкіна - Хакслі
Розберемо розвиток збудження на прикладі нервового волокна. Теорій, що пояснюють розвиток процесу збудження багато, але ми зупинимося на одній з останніх, експериментально найбільше підтвердженої. Це теорія Ходжкина - Хакслі, або теорія калієво - натрієвого насоса. Відповідно до цієї теорії в мембрані клітин, товщиною біля 75А (1А - 10-8 мм) є велика кількість пор і каналів діаметром 7 - 8 А. Пори несуть визначений електричний заряд, що полегшує або утруднює дифузію іонів через мембрану.
Мембрана більш проникна для катіонів, ніж для аніонів. Різні катіони також проходять через мембрану з неоднаковою швидкістю.
За допомогою мікроелектродної техніки встановлено, що у стані спокою електричний потенціал на мембрані позитивний, а усередині - негативний. Різниця потенціалів пояснюється іонним складом усередині і поза клітиною. Різниця в іонному складі пояснюється насамперед неоднаковим розподілом іонів натрію і калію - так у цитоплазмі нервових м'язових клітин концентрація калію майже в 40 разів вище, чим зовні, а натрію - у 12 разів вище зовні, чим усередині. Для простоти можна вважати, що іони натрію знаходяться зовні клітини, а іони калію - всередині. При цьому спеціальні системи активно переносять іони калію через мембрану усередину клітини і викидають іони натрію, що просочилися в результаті дифузії в клітину з неї.
Звичайно, цей опис будови і функції клітинної мембрани дуже спрощений, але він дає нам можливість роздивитися, як виникає і поширюється нервовий імпульс, тобто хвиля збудження.
Початковою реакцією клітини на подразнення є зміна проникності клітинної мембрани в місці подразнення. Іони натрію миттєво надходять усередину клітини, вносячи позитивний заряд.
У цей же час із клітини виходить незначна кількість іонів калію. За один імпульс клітина втрачає мільйонну частину всієї кількості калію. У результаті іонних переміщень наступає деполяризація - клітинний потенціал падає до нуля. Потім відбувається перезарядження мембрани - її реполяризація. Через якийсь час потенціал мембрани відновляється і клітина знову може реагувати на подразнення.
Надходження іонів натрію всередину клітини є причиною появи місцевого кругового електричного струму, що поширюється по поверхні мембрани. Всередині клітини круговий струм йде від збудженого місця до незбудженого, а зовні - від місця спочинку до збудженої ділянки. Проникаючи в сусідню незбуджену ділянку, місцевий струм відчиняє пори для натрію, створюючи тут новий імпульс. Зароджуючись у кожній такій точці нервового або м'язового волокна, імпульс поширюється на значну відстань.
Нервовий імпульс - складний електрохімічний процес. Для переміщення іонів калію і натрію проти градієнту концентрації необхідна енергія. Ця енергія утворюється в результаті розщеплення молекул АТФ.
Жива клітина при здійсненні своїх функцій безперервно генерує і підтримує електричні потенціали, тому універсальним способом з'ясування механізмів діяльності збудливих структур є електрофізіологічні дослідження. Основними іонами, які беруть участь в генерації електричних потенціалів клітин, є К+, Nа+ , Са2+ і Сl–. За умовами виникнення в живих структурах розрізняють мембранні потенціали спокою(МПС) та потенціали дії(ПД).
Мембранний потенціал спокою
Мембрана клітини електрично поляризована, і різницю потенціалів між зовнішньою і внутрішньою поверхнями клітинної мембрани називають мембранним потенціалом спокою(МПС). Для того щоб зрозуміти природу виникнення цього потенціалу, треба розглянути деякі властивості самої мембрани, а також склад поза- і внутрішньоклітинної рідин.
Клітинна мембрана має значний опір, який значною мірою залежить від стану її йонних каналів і ємність, яка створюється переважно її ліпідним матриксом, і є досить сталою. Так, у аксона кальмара питомий опір клітинної мембрани (Rм) становить у стані спокою 1000 Ом•см2, а ємність (См) — приблизно 1 мкФ/см2.
Існування МПС зумовлене передусім нерівномірною концентрацією іонів у поза- та внутрішньоклітинному просторі.
Другим важливим чинником, який має значення для створення МПС, є різна проникність клітинної мембрани для різних іонів. Клітинна мембрана легко проникна дляжиророзчинних речовин, молекули яких проходять крізь її ліпідний матрикс. Водорозчинні молекули великого діаметра, у тому числі й аніони органічних кислот, практично не проникають крізь мембрану У стані фізіологічного спокою клітинна мембрана проникна лише для К+, оскільки в ній є спеціальні калієві канали,що пропускають лише гідратований калій.
Коли К+ виходить з клітини, він затримується на зовнішній поверхні клітинної мембрани, створюючи її позитивний заряд. Великі аніони не можуть пройти з клітини крізь мембрану і накопичуються біля її внутрішньої поверхні, створюючи негативний потенціал. Так виникає трансмембранна різниця потенціалів,або МПС.
Цей потенціал протидіє подальшому виходу К+ за електрохімічним градієнтом.
Переміщення К+ із клітини назовні зумовлюється його концентраційним градієнтом, який здійснює осмотичну роботу (Аосм).
В наслідок МПС К+ частково повертається назад у клітину. При цьому здійснюється електрична робота(Аел).
. 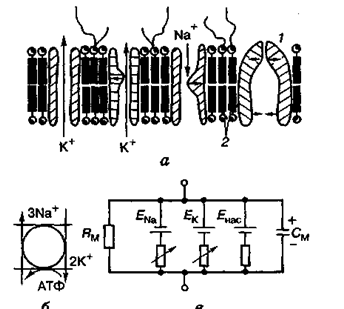
Мал. 1. Будова клітинної мембрани (о), її еквівалентна електрична схема (е) і схема активного перенесення йонів за рахунок енергії АТФ (б): / — іонний насос; 2 — ліпідний матрикс (білкові молекули, то утворюють іонні канали і насос, заштриховані); горизонтальними стрілками показано ворітні механізми, вертикальними — напрямок руху іонів; ЕNа, ЕК, Енас — потенціал натрію, калію, насоса; Rм, См — опір та ємність клітинної мембрани
Якщо вихід К+ з клітини переважає над надходженням, то поступово зростає Аелі дещо зменшується аосм. Внаслідок цього на клітинній мембрані виникає потенціал Е, за якого Аел=аосм для К+ і який називають калієвим рівноважним потенціалом(ЕК).
Різницю між поточним значенням МПС і Екназивають електрохімічним градієнтом для КК. Він є основною причиною пасивного рухуК+ крізь мембрану в природних умовах.
Якщо відношення внутрішньої і зовнішньої концентрації іонів калію
Слід зауважити, що на підтримання МПС витрачається дуже мало К+, основна їх частина повинна забезпечувати електронейтральністьвнутрішнього середовища клітини. Адже внутрішньоклітинні аніони – це переважно великі білкові молекули, які не можуть пройти крізь клітинну мембрану, і тому їх концентрація залишається сталою. Ці великі аніони мають бути зрівноважені відповідною кількістю катіонів. У зв'язку з тим, що вміст Nа+ всередині клітини дуже незначний, цю функцію виконують К+. Тому внутрішньоклітинна концентрація К+ має бути такою ж високою, як і концентрація великих аніонів, і такою ж стабільною.
МПС створює електричне поле, яке діє на макромолекули мембрани й надає їхнім зарядженим групам певну просторову орієнтацію. Електричне поле МПС також забезпечує функцію йонних каналів клітинної мембрани.
У стані спокою клітинна мембрана є добре проникною не лише для К+, вона певною мірою проникна також для інших іонів, зокрема Nа+, С1–, Са2+. Потоки цих іонів порушують рівновагу. Тої факт, що в умовах спокою Nа+ постійно надходить у клітину, а К+ виходить з неї, має дуже важливі наслідки, оскільки система не може бути фактично врівноважена, за допомогою звичайної дифузії і простого відновлення заряду на клітинній мембрані. Якщо немає інших процесів, то клітина дуже швидко накопичуватиме Nа+, втрачатиме К+, що зменшить їх трансмембранний градієнт. Отже, цей процес може призвести до різкого зниження МПС і загибелі клітини. Проте такого не відбувається, оскільки цьому протидіє натрій-калієвий насос.
Оскільки Nа+ сам не може вийти з клітини, а К+ — надійти в клітину проти своїх електрохімічних градієнтів, вони повинні активно переміщуватися крізь мембрану і на забезпечення цього транспорту має витрачатися енергія. Такий механізм дістав назву натрій-калієвого насоса.Він працює, споживаючи енергію АТФ, і його основним компонентом є фермент мембранна натрій-калій-АТФ-аза. У нормі АТФ потрапляє до насоса з мітохондрій аксоплазми. Для роботи насоса обов'язково мають бути зовні К+, а всередині — Nа+.
Енергія розщеплення однієї молекулі АТФ забезпечує виведення з клітині трьох іонів натрію і введення всередину клітини двох іонів калію. Це означає, що натрій-калієвий насос є електрогенним:він створює на клітинній мембрані додаткову різницю потенціалів Емембр, яка додається до рівня МПС.
Цей електрогенний додаток до нормального рівня МПС неоднаковий для різних клітин. Зазвичай у нервових клітин хребетних він незначний, а в гігантських нейронах молюсків і гладком'язових клітинах може становити до 25 % значенні МПС.
У формуванні МПС натрій-каліевий насос виконує дві функції:
1) підтримує трансмембранний градієнт концентрацій Nа+ і К+;
2) генерує певну різницю потенціалів, яка додається до потенціалу що створюється дифузією К+ за концентраційним градієнтом.
Важливим показником здатності іонів проходити крізь клітинну мембрану є її проникність. Значення проникності для іонів К+,Nа+ і Сl– (РК РNа, РСl) можна використовувати для обчислення МПС мембрани, проникної для всіх цих іонів, за рівнянням Гольдмана — Ходжкіна— Катца (рівняння постійного поля).
Потенціал дії
Потенціалом дії (ПД) називають швидке коливання МПС, що виникає під час збудження клітини.
В основі ПД лежать зміни електричної (іонної) провідності клітинної мембрани. Амплітуда і тривалість ПД окремої клітини мало залежать від сили подразнення, важливо лише, щоб ця сила досягла певного критичного значення — порогової сили подразнення. ПД, що виникає у місці подразнення, поширюється вздовж нервового чи м'язового волокна, не змінюючи своєї амплітуди. Наявність порогу подразнення і незалежність амплітуди ПД від сили подразнення підпорядковуються закону "все або нічого".
Мал. 2. Механізм виникнення двофазного потенціалу дії:
1 - деполяризація; 2 – реверсія (перезарядка мембран, овершут); 3 – реполяризація; 4 – слідова деполяризація; 5 – слідова гіперполяризація
У ПД розрізняють пік (верхівку), висхідну, низхідну фази і слідові потенціали. Встановлено, що під час висхідної фази ПД відбувається не просто зникнення трансмембранної різниці потенціалів (деполяризація мембрани),а виникає різниця потенціалів протилежного знака (овершут), внутрішня частина мембрани в ділянці генерації ПД стає на певний час зарядженою позитивно відносно зовнішньої частини мембрани, зарядженої негативно.
Під час низхідної фази ПД відновлюється початкове значення МПС (реполяризація мембрани).Тривалість піку ПД різних збудливих тканин варіює від 0,5 до 3 мс. Зміни поляризації мембрани, які спостерігаються після завершення високовольтної частини ПД (піку), називають слідовими потенціалами. Розрізняють два види слідових потенціалів — слідову деполяризацію(слідовий негативний потенціал) і слідову гіперполяризацію(слідовий позитивний потенціал), амплітуда яких не перевищує кількох мілівольт (5-10% рівня піку), а тривалість буває різною — від кількох мілісекунд до десятків секунд .
Іонний механізм генерації ПД.В основі генерації ПД лежать зміни проникності клітинної мембрани, що виникають послідовно за часом. У момент подразнення провідність мембрани для Nа+ зростає. Nа+ починає надходити в клітину, знижуючи внутрішній негативний заряд мембрани, тобто виникає деполяризація клітинної мембрани. При досягненні мембраною певного — критичного— рівня деполяризації виникає лавиноподібний потік Nа+ всередину клітини, що зумовлює появу висхідного коліна ПД. У цей момент gNа зростає у 100 разів порівняно зі станом спокою і стає у 20 разів більшої за gк. І якщо gNа збільшується далі, то заряд мембрани в місці подразнення змінюється — виникає реверсія знакаМП. Проте під час збудження позитивне значення ПД (овершут) не може бути білішим за натрієвий рівноважний потенціал,(ЕNа), який становить +65мВ. Звичайно овершут не перевищує + 30 мВ.
Натрієва провідність досягає максимуму менш ніж за 1 мс після початку ПД, а потім різко знижується, зникаючи зовсім протягом 0,5 мс, хоча деполяризація мембрани ще зберігається. Це явище називають натрієвою інактивацією,яка існує доти, доки триває деполяризація мембрани. Натрієва активаційна система відновлюється, коли МПС повертається до початкового рівня або навіть при певній гіперполяризації мембрани. На відміну від gNа, gк не інактивується і навіть зростає доти, доки існує деполяризація мембрани.
Дата добавления: 2018-06-27; просмотров: 126; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!