Мембранно-ионная теория биопотенциалов
Первым, кто предложил ионную теорию биоэлектрических явлений, был украинский физиолог, академик АН Украины Василий Юрьевич  Чаговец. В 1896 им была опубликована статья о применении теории электролитичической диссоциации для объяснения происхождения электрических явлений в живом организме (автор теории электрической диссоциации – шведский ученый Сванте Август Аррениус). Так была предпринята первая попытка объяснения токов покоя (т.н. демаркационных токов) в мышце, исходя из современных физико-химических представлений, и заложены основы ионной теории возбуждения.
Чаговец. В 1896 им была опубликована статья о применении теории электролитичической диссоциации для объяснения происхождения электрических явлений в живом организме (автор теории электрической диссоциации – шведский ученый Сванте Август Аррениус). Так была предпринята первая попытка объяснения токов покоя (т.н. демаркационных токов) в мышце, исходя из современных физико-химических представлений, и заложены основы ионной теории возбуждения.
В.Ю. Чаговец Академик Чаговец считал, что электрические токи в
живых тканях (токи покоя и токи действия) являются концентрационными, то есть, обусловлены накоплением в ткани продуктов обмена веществ. В 1906 он развил конденсаторную теорию электрических раздражения живых тканей. Идеи Чаговца получили дальнейшее развитие в трудах многих физиологов.
В Европе в эти же годы дугой ученый – немецкий физиолог, ученик Реймона, Юлиус Бернштейн также сосредоточил свои интересы в области нейробиологии и биофизики. В 1902 году он выдвинул мембранную теорию электрических потенциалов. Причиной возникновения биопотенциалов Бернштейн считал неодинаковое распределение ионов (заряженных частиц) по разные стороны клеточной мембраны. На внешней поверхности мембраны положительный заряд создается преимущественно катионами натрия. На внутренней поверхности концентрация катионов натрия очень низка. Для катионов калия ситуация складывается противоположная.  Отрицательный заряд внутренней поверхности клеточной мембраны создается за счет анионов радикальных групп крупных молекул органических соединений. Поры в мембране легко пропускают ионы калия, но для ионов натрия, как и для органических соединений по убеждению Бернштейна, транспорт практически невозможен. Потенциал покоя возникает из-за тенденции катионов калия диффундировать через клеточную мембрану изнутри наружу. Бернштейн также предположил, что
Отрицательный заряд внутренней поверхности клеточной мембраны создается за счет анионов радикальных групп крупных молекул органических соединений. Поры в мембране легко пропускают ионы калия, но для ионов натрия, как и для органических соединений по убеждению Бернштейна, транспорт практически невозможен. Потенциал покоя возникает из-за тенденции катионов калия диффундировать через клеточную мембрану изнутри наружу. Бернштейн также предположил, что
|
|
|
Ю. Бернштейн нервный импульс вызывается потенциалом действия
(электрической стимуляцией). При этом происходит изменение проницаемости мембраны, ионы с обеих сторон приходят в контакт и нейтрализуют потенциал покоя; после чего возвращается исходная проницаемость и, соответственно, потенциал покоя.
 Британский нейрофизиолог и биофизик Алан Ллойд Ходжкин предположил, что допущение Бернштейна по поводу отсутствия транспорта ионов натрия ошибочно. Многочисленные эксперименты (50-60гг. ХХв), проведенные им с коллегами Беркером и Шоу с использованием высокоточных методов на мембране гигантского аксона кальмара (освобожденного от протоплазмы) показали, что на самом деле измеряемый потенциал покоя всегда ниже, чем вычисленный теоретически по формуле Нернста. Это было прямым доказательством того, что кроме катионов калия существовал еще какой-то фактор, не учтенный теорией Бернштейна. В дальнейших испытаниях мембраны гигантского аксона
Британский нейрофизиолог и биофизик Алан Ллойд Ходжкин предположил, что допущение Бернштейна по поводу отсутствия транспорта ионов натрия ошибочно. Многочисленные эксперименты (50-60гг. ХХв), проведенные им с коллегами Беркером и Шоу с использованием высокоточных методов на мембране гигантского аксона кальмара (освобожденного от протоплазмы) показали, что на самом деле измеряемый потенциал покоя всегда ниже, чем вычисленный теоретически по формуле Нернста. Это было прямым доказательством того, что кроме катионов калия существовал еще какой-то фактор, не учтенный теорией Бернштейна. В дальнейших испытаниях мембраны гигантского аксона
|
|
|
кальмара (А. Ходжкин и Бернард Катц), было
А. Ходжкин установлено, что потенциал покоя, измеряемый в опыте,
оказывается ближе всего к теоретическому в одном особом случае: если из внеклеточной среды полностью вымывали катионы натрия. Предположение, что ионы натрия могут проходить через мембрану гигантского аксона, было прямо доказано в опытах с радиоактивным изотопом натрия. Побочным результатом этих экспериментов было открытие яда, тетродотоксина (содержится в рыбе фугу), блокирующего движение натрия через мембрану. При добавлении тетродотоксина в морскую воду, окружающую гигантский аксон, натрий перестает идти через мембрану, потенциал покоя начинает нарастать и достигает значения, соответствующего формуле Нернста. Так было показано, что мембранный потенциал является результатом движения по электрохимическому градиенту двух встречных ионных потоков – калия и натрия.
|
|
|
Впоследствии был установлен факт, доказывающий, что поддержание постоянного уровня потенциала покоя клетки зависит не только от трансмембранных ионных градиентов калия, натрия, но и хлора. Были открыты ионные насосы, использующие энергию АТФ, поддерживающие постоянство трансмембранных ионных градиентов.
В 1963 году А́лан Ллойд Хо́джкин совместно с австралийским нейрофизиологом Джоном Эклсом и английским нейрофизиологом и биофизиком Эндрю Хаксли были удостоены нобелевской премии «за открытия, касающиеся ионных механизмов возбуждения и торможения в периферических и центральных участках нервных клеток». Так было совершено открытие потенциала действия, объясняющего природу возникновения нервного импульса.
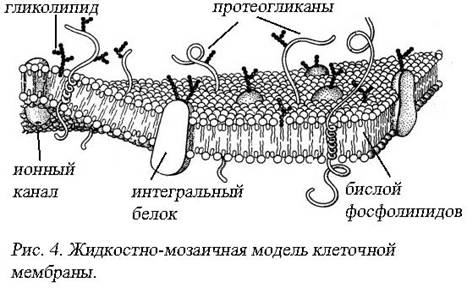
Важно отметить, что успешному развитию электрофизиологии в середине ХХ века способствовали успехи клеточной биологии. Так, в 1935 году появляется унитарная модель строения биомембран, предложенная Дэниэли Доусоном. Однако быстрое развитие физических и химических методов исследования, дало возможность к 70-м годам предложить новую модель строения биологических мембран – жидкостно-мозаичную (С. Сингер, Г. Никольсон, 1972). Белково-кристаллическая модель (Г. Вандеркой, Д. Грин, 1970) отличается от жидкостно-мозаичной модели фактически лишь постулированием существования в мембране жесткой белковой структуры, возникающей в результате дальнодействующих белок-белковых связей. В настоящее время получили наибольшее распространение различные варианты жидкостно-мозаичной модели (рис. 4).
|
|
|
Основные положения, сформированные мембранно-ионной теорией биопотенциалов:
1. Мембрана живой клетки всегда поляризована. Состояние поляризации проявляется в том, что на внешней поверхности клеточной мембраны преобладают преимущественно катионы, на внутренней – преимущественно анионы.
2. Мембрана живой клетки обладает избирательной проницаемостью. Это свойство связано как с особенностями строения трансмембранных белков-переносчиков (ионных каналов и ионных насосов), так и с физико-химическими параметрами электролитов.
3. Мембрана электровозбудимой клетки при действии стимула способна быстро менять свою проницаемость для определенного вида катионов, что вызывает распространяющиеся процессы деполяризации. Подведем итог, только в ХХ веке появились экспериментальные доказательства того, что «животное электричество», проявляющее себя как потенциал покоя и потенциал действия, обусловлено движением ионов натрия, калия, кальция, хлора через специальные сложно организованные ионные каналы в мембране, а также обусловлены диффузией ионов в межклеточной и внутриклеточной средах, представляющих собой растворы электролитов.
 |
Дата добавления: 2018-06-27; просмотров: 124; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!