Сравнительная характеристика активных состояний возбудимых тканей
| Состояние активности Основные характеристики | Возбуждение | Торможение |
| Неспецифические признаки | Изменение проницаемости клеточной мембраны. Выраженное изменение уровня обменных процессов: потребление О2 и выделение СО2, изменение вязкости протоплазмы, температуры и т.д. | |
| Специфические признаки | Нервная клетка проводит нервный импульс, мышца сокращается | Блокируется проведение нервного импульса, мышца не сокращается |
| Движение ионов через клеточную мембрану | Повышается проницаемость для Na+ | Повышается проницаемость для К+ и Cl- |
| Изменение заряда клеточной мембраны | Деполяризация – инверсия поляризации | Гиперполяризация – усиление поляризации |
| Затраты энергии | Энергоемкий процесс, энергия тратится как на совершение работы, так и на поддержание гомеостаза | Энергосберегающий процесс, работа не совершается. Энергия тратится только на поддержание гомеостаза |
Таблица 2
Сравнительная характеристика местного и распространяющегося возбуждения
| Вид возбуж- дения Харак- теристики | Местное возбуждение (локальный ответ, локальный потенциал) | Распространяющееся возбуждение (потенциал действия, нервный импульс) |
| Происхождение | Наиболее древний вид возбуждения, современное определение – раздражимость. Встречается у низших форм организмов и в низковозбудимых тканях (напр. соединительной) | Присуще высокоорганизованным возбудимым тканям |
| Сила раздражителя | Подпороговый | Пороговый и надпороговый |
| Распространение | С декрементом (затуханием) на 1-2мм от места действия стимула | Без декремента по всей длине нервного волокна |
| Зависимость от величины силы раздражителя | Градуально – изменение заряда клеточной мембраны пропорционально силе подпорогового раздражителя (подчиняется закону «силы») | Не градуально – изменение заряда клеточной мембраны не зависит от силы раздражителя, если его величина превысила пороговое значение (подчиняется закону «все или ничего») |
| Скорость развития возбуждения | Нет латентного (скрытого) периода. Возбуждение возникает сразу же при действии раздражителя. | Есть латентный период – между моментом нанесения раздражения и видимой ответной реакцией проходит некоторое время |
| Изменение возбудимости | Нет рефрактерного периода, есть кратковременное повышение возбудимости | Есть рефрактерный период |
| Амплитуда | 10-40мВ | 80-130мВ |
| Способность суммироваться | Пространственная и временная суммация | Нет суммации |

Биоэлектрические явления
Свои первые предположения о существовании электрических явлений у живых организмов Луиджи Гальвани высказал в небольшой научной публикации «О влиянии электрических сил на движение мускулов животных» (Comment. Bonon., VII, 1791). Гальвани обнаружил электрический ток в цепи, составленной из металлов и лягушечьих мышц, установил ряд его свойств и правильно определил на опыте влияние на этот ток металлов и тел животных.
Луиджи Гальвани Однако он ошибся в определении
действительного источника этого электричества. Гальвани предполагал, что у всех животных электричество рождается в мозгу и распространяется отсюда по нервам к мышцам. Считая металлы в этих явлениях только проводниками, он употреблял медь или серебро наряду с железом только из-за их большей проводимости. Гальвани не был физиком и не мог предположить, что контакты разных металлов могут служить источником электрических возмущений. Впоследствии эти опыты удавались ему и без применения металлов, когда он даже препарирование мышц производил стеклянными ножами (рис.2). Действительно открытое им и не признанное его противниками животное или физиологическое электричество помешало ему заметить, что в большинстве прежних его опытов наблюденные им явления вызывались не этим электричеством, а другим, более сильным металлическим электричеством.
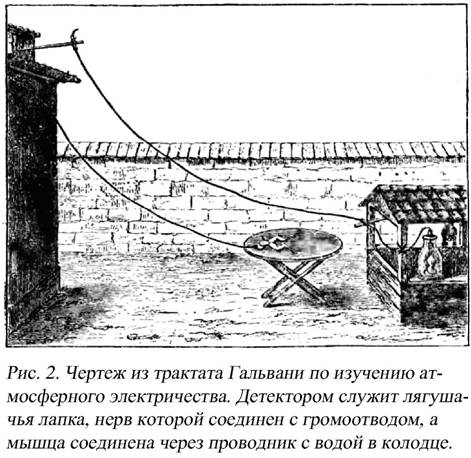
Эксперименты вызвали большой резонанс в научном обществе. Утверждение о естественном присутствии электричества в животных органах опровергалось Алессандро Вольтой. Многократно повторяя эксперименты Гальвани, Вольта нашел два недостатка в методической части. Во-первых, электрический ток в опытах Гальвани вызывал не непосредственно сокращение мышцы, а приводил в состояние возбуждения нерв, который вызывал рефлекс мышечного сокращения. Во-вторых, на основании множества опытов Вольта пришел к убеждению, что два разнородных металла могут быть источником электричества, а живые ткани – только проводниками электрического тока. Так был изобретен первый гальванический элемент, который по иронии судьбы получил название «элемент Вольта» или «вольтов столб».
Тем не менее, правильная оценка наблюдений Гальвани стала возможной лишь после изобретения достаточно чувствительных электроизмерительных приборов – гальванометров. Итальянский физик Карло Маттеуччи в 1837 году доказал наличие разности электрических потенциалов между поврежденной и неповрежденной частями мышцы. Согласно экспериментальным данным электрический ток течёт от её неповрежденной поверхности к поперечному разрезу. Также Маттеуччи обнаружил, что мышца при её сокращении создаёт электрический ток, достаточный для раздражения другого нервно-мышечного соединения.
 Но ответить на вопрос «откуда же берется животное электричество?» удалось другому замечательному ученому – немецкому физиологу, философу Эмилю Генриху Дюбуа-Реймону. Еще в студенческие годы на втором курсе Берлинского университета он остановил свой выбор на физиологии. Посвятив исследованиям влияния электрического тока
Но ответить на вопрос «откуда же берется животное электричество?» удалось другому замечательному ученому – немецкому физиологу, философу Эмилю Генриху Дюбуа-Реймону. Еще в студенческие годы на втором курсе Берлинского университета он остановил свой выбор на физиологии. Посвятив исследованиям влияния электрического тока
на нервы несколько лет, Дюбуа-Реймон, опубликовал в
К. Маттеуччи 1843 году труд, который ознаменовал начало
 современной электрофизиологии – "Предварительный очерк исследования о так называемом лягушачьем токе и об электромоторных рыбах". Всю свою творческую жизнь ученый посвятил электрофизиологии, он организовал мастерскую, в которой изобретали и собирали приборы для изучения «животного электричества» Дюбуа-Реймон является автором молекулярной теории биопотенциалов: биоэлектрические явления в тканях – это разность потенциалов, которая возникает в тканях в процессе нормальной жизнедеятельности.
современной электрофизиологии – "Предварительный очерк исследования о так называемом лягушачьем токе и об электромоторных рыбах". Всю свою творческую жизнь ученый посвятил электрофизиологии, он организовал мастерскую, в которой изобретали и собирали приборы для изучения «животного электричества» Дюбуа-Реймон является автором молекулярной теории биопотенциалов: биоэлектрические явления в тканях – это разность потенциалов, которая возникает в тканях в процессе нормальной жизнедеятельности.
Эти явления удалось зарегистрировать,
используя трансмембранный способ регистрации
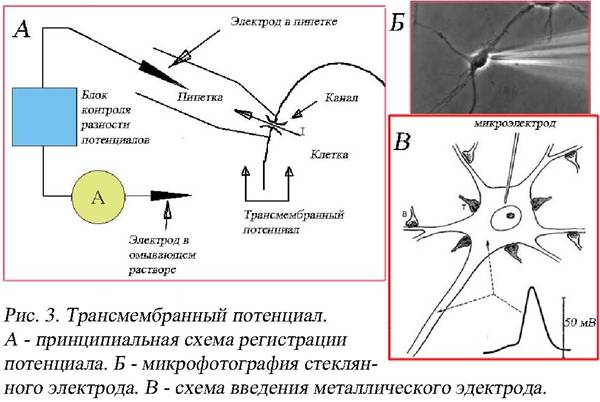
Э. Дюбуа-Реймон (рис. 3) при помощи микроэлектродной техники. Один
электрод (он может быть металлическим или стеклянным) вводят внутрь клетки, а другой, являющийся индифферентным электродом, помещают на поверхности клетки или чаще в межклеточном пространстве. Оба микроэлектрода связаны очень чувствительным осциллографом, который может многократно усиливать и регистрировать электрический потенциал, отведенный от возбудимой клетки. Полученная кривая называется клеточной трансмембранной электрограммой. Для клеток нервной системы микроэлектродную технику стали применять в 1951 году в лаборатории австралийского нейрофизиолога Дж. Экклса. Трансмембранный потенциал – это потенциальная разница между внутренней и наружной сторонами клетки. Таким способом регистрируется мембранный потенциал покоя и потенциал действия.
Замечательным этапом в развитии электрофизиологии стало открытие, сделанное в 1875 году английским хирургом, физиологом Ричардом Кэтоном и русским физиологом Василием Яковлевичем Данилевским. Они независимо друг от друга пришли к выводу о генерации мозгом электрической активности – биотоков мозга.
 |
Большой вклад в теорию биоэлектрических явлений внесли труды  немецкого химика Вальтера Германа Нернста. В 1888-89 гг. он изучал поведение электролитов при пропускании электрического тока и открыл фундаментальный закон, известный как уравнение Нернста. Закон устанавливает зависимость между электродвижущей силой (разностью потенциалов) и ионной концентрацией. Вальтер Нернст научно обосновал природу равновесия между диффузией и другими видами транспорта через мембрану. До Нернста
немецкого химика Вальтера Германа Нернста. В 1888-89 гг. он изучал поведение электролитов при пропускании электрического тока и открыл фундаментальный закон, известный как уравнение Нернста. Закон устанавливает зависимость между электродвижущей силой (разностью потенциалов) и ионной концентрацией. Вальтер Нернст научно обосновал природу равновесия между диффузией и другими видами транспорта через мембрану. До Нернста
считалось, что диффузия различных ионов через
В. Нернст мембранные каналы должна в итоге привести к
выравниванию концентраций ионов между вне- и внутриклеточной средой. Однако эти изменения сохраняются, пока клетка жизнеспособна (табл. 3). Уравнение Нернста показывает, что величина мембранного потенциала, является результатом электрохимического взаимодействия, этот закон связывает термодинамику с электрохимической теорией в области решения проблем, касающихся сильно разбавленных растворов:
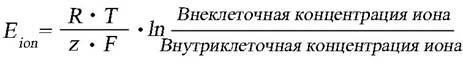
где R – газовая постоянная, T – абсолютная температура,
z – валентность иона (отрицательная для анионов), F – число Фарадея.
Удивительным оказался тот факт, что если подставить в уравнение константы, то при температуре тела (Т=310К0, или 36,6 С0) равновесный потенциал для ионов калия составит ‒97мВ. И действительно было обнаружено, что мышечные клетки млекопитающих имеют мембранный потенциал равный ‒90мВ. В зависимости от условий и относительных концентраций ионов величина мембранного потенциала может колебаться от ‒40 до ‒120мВ.
Благодаря этой работе 25-летний Нернст завоевал всемирное признание. Закон распределения Нернста сыграл большую роль для медицины и биологии, поскольку позволил исследовать распределение веществ в различных частях живого организма.
Дата добавления: 2018-06-27; просмотров: 207; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!