Синтез глюкозы в результате глюконеогенеза энергозависим При синтезе глюкозы из лактата расходуется энергия 4 АТФ и 2 ГТФ
лактат + лактат → 1 молекула глюкозы
• пируваткарбоксилаза – 2 АТФ,
• фосфоенолпируваткарбоксикиназа – 2 ГТФ
• на этапе обратимой киназной реакции гликолиза при глюконеогенезе идёт уже
не образование, а расход АТФ – 2 АТФ
Таким образом – при низком содержании углеводов в пище, голодании, длительной физической работе – организм должен иметьвозможность получить глюкозу. Это достигается процессом глюконеогенеза. Кроме получения глюкозы, глюконеогенез обеспечивает и уборку "шлаков" – лактата, образованного при мышечной работе и в эритроцитах, и глицерола, являюще-
гося продуктом липолиза в жировой ткани
Глюконеогенез – синтез глюкозы
de novo из неуглеводных компонентов.
Протекает в печени и ≈10% в почках.
• Предшественники для
глюконеогенеза
• лактат (главный),
• глицерол (второй),
• аминокислоты (третий) – в условиях
длительного голодания.
• Физиологическое значение.
Места поступления субстратов (предшественников) для
глюконеогенеза


аланин
Биологический смысл глюкозо-аланинового цикла – транспорт аминного азота из мышц и предотвращение лактоацидоза

20. Пути образования и использования пировиноградной кислоты. Окислительное декарбоксилирование пирувата: последовательность реакций, связь с дыхательной цепью, регуляция, энергетический эффект. Метаболизм пирувата в реакциях трансаминирования и глюконеогенеза, биологическое значение этих процессов. Обмен пирувата и баланс витаминов. Характеристика тиамина и пантотеновой кислоты, их роль. Особенности метаболизма пирувата микрофлорой полости рта.
Пируват — важное химическое соединение в биохимии. Он является конечным продуктомметаболизма глюкозы в процессе гликолиза. Одна молекула глюкозы превращается при этом в две молекулы пировиноградной кислоты. Дальнейший метаболизм пировиноградной кислоты возможен двумя путями — аэробным и анаэробным. В условиях достаточного поступления кислорода, пировиноградная кислота превращается в ацетил-кофермент А, являющийся основным субстратом для серии реакций, известных как цикл Кребса, или дыхательный цикл, цикл трикарбоновых кислот. Пируват также может быть превращён в анаплеротической реакции в оксалоацетат. Если кислорода недостаточно, пировиноградная кислота подвергается анаэробному расщеплению с образованием молочной кислоты у животных и этанол[1] у растений.
ОКИСЛИТЕЛЬНОЕ ДЕКАРБОКСИЛИРОВАНИЕ ПИРУВАТА
1-ю реакцию катализирует фермент ПИРУВАТДЕКАРБОКСИЛАЗА (Е1).


|
2-ю и 3-ю реакцию процесса катализирует фермент АЦИЛТРАНСФЕРАЗА (Е2).
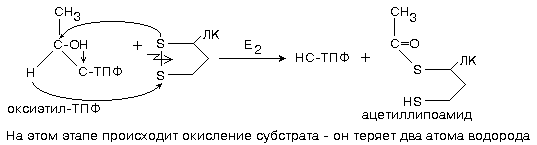
3 этап - продолжает работать фермент ацилтрансфераза.

Второй фермент данного комплекса является не только ацетилтрансферазой, но и окислительным ферментом одновременно (дегидрогенизирующая ацетилтрансфераза).
Энергия этого окисления аккумулируется в виде макроэргической связи активной формы уксусной кислоты (ацетил-КоА).
4 этап катализируется ферментом ДИГИДРОЛИПОИЛДЕГИДРОГЕНАЗОЙ.

В итоге можем записать суммарное уравнение:

Аналогично действует ферментный комплекс для окислительного декарбоксилирования a-кетоглутарата. Но названия двух из трех ферментов другие:
1-й фермент - декарбоксилаза a-кетоглутаровой кислоты
2-й фермент - сукцинилтрансфераза (переносит остаток янтарной кислоты)
Суммарное уравнение для этого комплекса:


В результате действия этих двух комплексов, каждый из двух приведенных субстратов (ПВК и a-КГ) - теряют СО2, два протона и два электрона, которые передаются на кислород с образованием Н2О и параллельно происходит фосфорилирование трех молекул АТФ. Сами субстраты превращаются в итоге в активную форму соответствующей кислоты.
Макроэргическая связь в молекулах этих активных форм может легко трансформироваться в макроэргическую связь молекулы АТФ. Например, для молекулы сукцинил-КоА:

Такой способ образования АТФ, когда нет сопряжения с процессами окисления, называется СУБСТРАТНЫМ ФОСФОРИЛИРОВАНИЕМ (при этом используется макроэргическая связь в молекуле какого-нибудь субстрата).
ЭНЕРГЕТИЧЕСКИЙ ИТОГ УДЛИНЕННОЙ ЦЕПИ.
Образуются 3 молекулы АТФ путем окислительного фосфорилирования. Продукты окислительного декарбоксилирования (ацетил-КоА и сукцинил-КоА) содержат макроэргическую связь. Энергия макроэргической связи сукцинил-КоА используется двумя способами.
1) Субстратное фосфорилирование в ЦТК (так расходуется большая часть этого вещества).
2) Реакции синтеза гема (меньшая часть сукцинил-КоА).
Энергия макроэргической связи Ацетил-КоА используется только в реакциях синтеза:
а) Синтез лимонной кислоты (цитрата).
б) Синтез кетоновых тел
в) Синтез жирных кислот
г) Синтез холестерина
Среди субстратов МтО почти нет веществ, которые организм получает с пищей - только глутаминовая кислота.
Субстраты МтО - в основном органические кислоты, которые образуются в организме в процессе катаболизма. В процессе катаболизма из большого количества разнообразных пищевых веществ образуется всего 2 вида общих метаболитов: Ацетил-КоА и ПВК.
Трансаминирование.При добавлении к гомогенату мышц глутаминовой и пиро-виноградной кислот образуются α-кетоглутаровая кислота и аланин без промежуточного свободного аммиака; добавление аланина и α-кетоглу-таровой кислоты приводило к образованию соответственно пировиноград-ной и глутаминовой кислот. Иными словами, предшественниками глюкозы вглюконеогенезе может быть пируват или любое соединение, превращающееся в процессе катаболизма в пируват или один из промежуточных продуктов цикла трикарбоновых кислот.
У позвоночных наиболее интенсивно глюконеогенез протекает в клетках печени и почек (в корковом веществе).
Большинство стадий глюконеогенеза представляет собой обращение реакции гликолиза. Только 3 реакциигликолиза (гексокиназная, фосфо-фруктокиназная и пируваткиназная) необратимы, поэтому в процесс глю-конеогенеза на 3 этапах используются другие ферменты. Рассмотрим путь синтеза глюкозы из пирувата.

Глюконеогенез – синтез глюкозы из неуглеводных продуктов. Такими продуктами или метаболитамиявляются в первую очередь молочная и пи-ровиноградная кислоты
Образование фосфоенолпирувата из пирувата.


ТИАМИН (витамин Bt), водорастворимый витамин группы В. Существует в виде солей, важнейшая из к-рых тиамин-гидрохлорид Соли тиамина хорошо раств. вводе, хуже в спиртах, не раств. в эфире, хлороформе, бензоле и ацетоне. 
Потребность человека в тиамине составляет. 0,5 мг на 1000 ккал (4186 кДж) суточного рациона или от 1,5 до 2,5 мг/сут в зависимости от энергозатрат. Тиамин широко распространен в живой природе. Синтезируется растениями и мн. микроорганизмами. Животные и человек не синтезируют тиамин и должны получать его с пищей. ТДФ-зависимая пируватдегидрогеназа принимает участие в окислит. декарбоксилировании пировиноградной к-ты (пирувата) с образованием ацетилкофермента А. Недостаток тиамина в организме ведет к накоплению в крови и тканях недоокисленных продуктов обмена в-в, что, в свою очередь, приводит к патофизиол. и патоморфологич. изменениям, создающим картину В1-авитаминоза, одной из форм к-рого является болезнь бери-бери (полиневрит, сердечно-сосудистые расстройства, отеки и др.).
Дата добавления: 2015-12-20; просмотров: 35; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!