Так, а кто из рекордсменов у нас есть в ранге морских Динозавров ? 2 страница
История изучения[править | править вики-текст]
Основная статья: История изучения динозавров
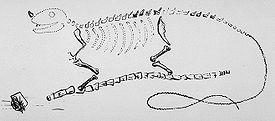
Реконструкция игуанодона Г. Мантелла — первая в истории реконструкция динозавра
Гигантские кости, находимые иногда в земле, в античности считали останками героев эпохи Троянской войны, в Средние века и вплоть до XIX в. — останками исполинов, о которых упоминается в Библии и которые погибли во время всемирного потопа; на Дальнем Востоке их считали костями драконов и приписывали им целебные свойства.
В 1824 году президент Королевского геологического обществаУильям Баклэнд выступил с докладом о находке, сделанной в 1815 году в юрских сланцах Стоунзфилда (графство Оксфордшир) и состоящей из нескольких костей и фрагмента «допотопного» животного. Прибегнув к помощи видного специалиста по сравнительной анатомииЖоржа Кювье, Баклэнд классифицировал находку как останки гигантской хищной ящерицы (лат. sauria) и, соответственно, назвал её мегалозавром — «огромным ящером».
В 1826 году Гидеон Мантелл, хирург из Льюиса (графство Суссекс), действительный член Линнеевского общества, аналогичным образом представил в Геологическом обществе найденные им зубы ранее неизвестного вида, которому он дал название игуанодон (букв. «игуанозубый») за сходство зуба с зубом ящерицы игуаны. Он же в 1833 году описал гилеозаврa — представителя панцирных ящеров анкилозавров.

Челюсть мегалозавра, иллюстрация из статьи Баклэнда
|
|
|

Мезозойская фауна в представлениях середины XIX в. На переднем плане изображены мегалозавры, реконструированные Р. Оуэном как четвероногие существа

Статуя мегалозавра в саду Хрустального дворца
В 1842 году английский биолог Ричард Оуэн, констатировав несомненное сходство между этими тремя видами и их отличие от современных рептилий, выделил их в особый подотряд, назвав его Dinosauria («ужасные ящеры»)[9].
Открытие в 1858 году в США хорошо сохранившегося скелета гадрозавра опрокинуло представление о динозаврах как о четвероногих животных, показав, что динозавры могли ходить на двух ногах. В последующие несколько десятилетий были открыты представители большинства основных групп динозавров; важная заслуга в этом принадлежит американским палеонтологам Гофониилу Маршу и Эдварду Копу, которые открыли и описали в общей сложности 142 новых вида, включая апатозавра и бронтозавра (впоследствии их отнесли к одному роду), диплодока и стегозавра, моноклона, трицератопса и др. Накопление материала привело к разделению динозавров на семейства птицетазовых и ящеротазовых (1887).
В рамках первой половины двадцатого века бо́льшая часть научного сообщества ошибочно полагала, что динозавры были громоздкими, вялыми животными. Большинство исследований, проводимых с 1970 года, однако, указали, что динозавры были активными животными с повышенным метаболизмом и многочисленными особенностями для социального взаимодействия.
|
|
|
В 1964 году находка дейнониха произвела новую научную революцию, так как по строению динозавра было ясно, что он передвигался относительно быстро, из чего следовал вывод, что он был теплокровным. Идея теплокровности заставляла пересмотреть старые представления не только о физиологии, но и о поведении динозавров, чему появились подтверждения в 1979 году, когда были получены доказательства родительского инстинкта и социального поведения ящеров (высиживание, защита и выкармливание детёнышей). Наконец, сравнение верхних конечностей дейнониха с крылом птицы заставило предполагать их близость и происхождение птиц от динозавров (или даже вообще принадлежность к этому надотряду), чему доказательством стало впоследствии открытие следов оперения у ряда динозавров. В 2005 году учёным удалось выделить коллаген из сохранившихся мягких тканей тираннозавра[10] и использовать его химический состав как ещё одно доказательство родства динозавров с современными птицами[11].
|
|
|
Настоящее время[править | править вики-текст]
Ряд учёных утверждает, что около трети описанных видов динозавров являются несуществовавшими. За ранее неизвестных ящеров учёные принимали уже описанных динозавров на разных стадиях развития. Проведённое масштабное исследование другими учёными показало, что почти 50 % всех видов динозавров были названы неправильно[12].
В настоящее время наблюдаются две противоположно направленные тенденции в систематике динозавров. В то время как одни палеонтологи продолжают дробление существующих таксонов, выделяя по тем или иным признакам новые рода и виды из уже существующих, другие их коллеги ставят под сомнение корректность ранее описанных видов. Так в 2007 году Джек Хорнер (англ. Jack Horner) опубликовал статью, в которой высказывал, что Dracorex hogwartsia, Stygimoloch spinifer и Pachycephalosaurus wyomingensis являются тремя разными возрастными стадиями одного вида. В 2009 году Джек Хорнер при изучении черепа Nanotyrannus заключил, что этот динозавр на самом деле является молодым Tyrannosaurus. В 2010 году в издании Journal of Vertebrate Paleontology вышла статья палеонтологов из Государственного университета Монтаны (Montana State University), где также говорится, что Triceratops и Torosaurus — различные стадии роста особей одного вида. В том же году другие учёные из Йельского университета описали новый род динозавров — Mojoceratops, кости которого раньше причисляли к Chasmosaurus[13].
|
|
|
Происхождение и эволюция[править | править вики-текст]
Предки[править | править вики-текст]
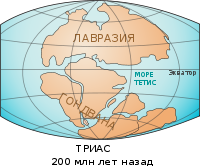
География земли в триасовом периоде
Рептилии обитали на Земле ещё до появления динозавров. Лапы у них были расположены по бокам туловища, как у большинства современных ящериц. Около 300 млн лет назад во время каменноугольного периода произошло глобальное потепление, что по мнению учёных уничтожило тропические леса и вызвало эволюционный взрыв среди рептилий. Каждая популяция оказалась запертой в небольшой области обитания, и каждая начала развиваться по-своему, что и привело к увеличению разнообразия[14].
Затем появились предки динозавров — архозавры («господствующие ящеры»). Появление архозавров шло параллельно с массовым (но не полным) вымиранием терапсид на границе перми и триаса (см. Массовое пермское вымирание). В начале триасового периода возникло множество новых видов.
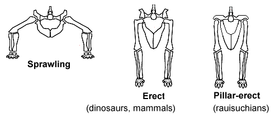
Изначальное положение задних конечностей у рептилий и два способа выпрямления — у динозавров и млекопитающих и у равизухий
Важнейшие продвинутые признаки ранних динозавров, которые отсутствовали у большинства текодонтов, были связаны с положением тела и характером локомоции. Задние конечности становятся практически вертикальными, а их длина по сравнению с более короткими передними конечностями позволяет делать выводы о хождении на двух ногах (бипедальности)[15].
Высшие формы терапсид по своей организации были очень близки к однопроходным млекопитающим и, по некоторым предположениям[16], имели молочные железы и шерсть. Внезапный упадок тероморфов и расцвет «настоящих ящеров» — одна из самых интригующих загадок палеонтологии[17].
Динозавры отклонились от своих предков архозавров спустя примерно 20 миллионов лет после массового пермского вымирания, которое уничтожило приблизительно 95 % всей жизни на Земле[18]. На сегодняшний день останки самых древних динозавров имеют возраст около 230 миллионов лет. Однако в 2010 году группа американских, немецких и южноафриканских палеонтологов представила в научном мире совместную работу, согласно с которой первые динозавры населяли Землю на несколько миллионов лет раньше, нежели было принято считать. Свои выводы они сделали, проанализировав останки новой динозаврообразной рептилии Asilisaurus kongwу, которая является родственным динозаврам видом, жившим 245—240 млн лет назад. Учёные говорят, что уже в это время различия между Dinosauriformes и самыми ранними динозаврами были существенными, следовательно, общий предок динозавров отделился от группы Ornithodira существенно раньше, чем считалось прежде[19].
По одной из гипотез, оба отряда динозавров впервые появились на территории современной Южной Америки и затем распространились по всей Пангее в течение триасового периода с 230 до 199 миллионов лет назад[20][21].
Первые динозавры[править | править вики-текст]

Скелеты эрреразавра и эораптора
Одним из самых ранних известных на сегодняшний день динозавров считается ставрикозавр, живший 228 млн лет назад, частичный скелет которого был обнаружен в отложениях среднего триасового периода на территории современной Бразилии. Большая берцовая кость длиннее бедренной, что характерно для бегающих животных, кости конечностей полые[22]. В 1963 году был описан эрреразавр, обнаруженный в отложениях позднего триаса на северо-западе Аргентины. Из-за фрагментарности останков точная его классификация была не определена. Лишь после того как в 1988 году был обнаружен более полный скелет с черепом, он в основном классифицировался как примитивный теропод[23]. Обнаруженный на территории современной Мексики вид Tawa hallae подтвердил связь эрреразавра с более поздними тероподами (хищными динозаврами)[24][25]. Эрреразавр достигал длины до 4-х метров[22] и веса 200—250 килограммов[23]. Череп длинный и узкий, не имеет признаков характерных для более поздних теропод[23], строение задних конечностей также характерно для ранних динозавров[22]. Через четыре года после открытия эрреразавра, в Аргентине был найден скелет ещё одного древнего динозавра — эораптора[26], жившего 228—235 млн лет назад. Внешне он был похож на других рептилий, но особенности костей и зубов позволяют отнести его к примитивным тероподам[27]. Палеонтологи полагают, что эораптор напоминает общего предка всех динозавров[28]. Если эти предположения верны, то основываясь на его чертах, можно говорить, что первые динозавры были маленькими, двуногими хищниками[26]. Открытия примитивных, подобных динозаврам орнитодиров, таких как маразух и Lagerpeton , поддерживают эти предположения; анализ окаменелостей данных рептилий действительно говорит об их малых размерах и хождении на двух ногах.
Самой первой известной группой птицетазовых динозавров являются Fabrosauridae (фаброзавриды). Самый ранний из них — пизанозавр (Pisanosaurus), обнаруженный в породах верхнего триаса на территории современной Аргентины. Лучше всего из фаброзавридов изучен род лесотозавр из юрских отложений Южной Африки. Как и все другие фаброзавриды он достигал в длину около 1 метра. Строение задних конечностей характерно для бипедальных животных, большая берцовая кость значительно длиннее бедренной. Некоторые части скелета сходны с таковыми у ранних теропод, но тазовые кости и строение черепа и зубов уже имеют признаки птицетазовых динозавров[22].
На самых ранних этапах эволюции динозавров земная среда была заселена различными видами архозавров и терапсидов, такими как этозавры, цинодонты, дицинодонты, орнитозухиды, ринхозавры и другие. По всей видимости, в конце триаса динозавры выдержали долгую конкуренцию с данными животными, прежде чем победили и достигли расцвета. Некоторые учёные, однако, полагают, что господствующее положение динозавров было получено не столько превосходством над другими животными, сколько случайностью. В карнийском веке динозавры не показывают образец устойчивого увеличения в разнообразии и числе, если бы они, конкурируя, занимали ниши других групп животных. Вместо этого они были очень редки, составляя только 1—2 % от общей животной фауны. В норийском веке, после исчезновения нескольких других групп, они стали существенными компонентами фауны, представляя уже 50—90 % живых существ[29]. Около 200 миллионов лет назад произошло так называемое триасовое вымирание. Это событие окончательно освободило экологические ниши, открыв возможность развиться, начиная с юрского периода, таким группам, как крокодиломорфы, птерозавры, млекопитающие, черепахи и динозавры[30].
Диверсификация в меловом периоде[править | править вики-текст]
Статистические исследования, основанные на имеющихся палеонтологических материалах, предполагают, что разнообразие динозавров, то есть число разновидностей, увеличилось в позднем меловом периоде.
Однако в июле 2008 года Грэм Т. Ллойд и его коллеги выдвинули утверждение, что это очевидное разнообразие является иллюзией, вызванной тем, что породы позднего мелового периода изучены в большей степени, чем породы юрского и триасового периодов. Вместо этого они написали статью о том, что в позднем меловом периоде существенной диверсификации подверглись только две группы динозавров — гадрозавры и цератопсы. В среднем меловом периоде цветущие покрытосеменные растения стали большей частью наземных экосистем, в то время как ранее доминировали голосеменные, такие как хвойные породы. Копролиты (окаменелый навоз) динозавров указывают на то, что в то время как некоторые ели покрытосеменные растения, большинство травоядных динозавров питалось, главным образом, голосеменными растениями. В это же самое время травоядные насекомые и млекопитающие развивались очень быстро и разносторонне, чтобы использовать в своих интересах новый тип пищи растительного происхождения. Ящерицы, змеи, крокодилы и птицы также разносторонне развивались в то же самое время. Ллойд и его коллеги предположили, что отказ динозавров разносторонне развиться в плане изменения экосистемы обрекал их на исчезновение[31].
Классификация[править | править вики-текст]
Динозавры относятся к группе архозавров, или «господствующих рептилий». К этой группе также относятся крокодилы, птерозавры и текодонты.
В филогенетической систематике динозавры обычно определяются как группа, состоящая из «трицератопсов, неорнитов (современных птиц), их общих предков и всех потомков»[30]. Также некоторыми учёными выдвигается предложение, чтобы динозавры были определены относительно общего предка мегалозавра и игуанодона, так как они были двумя из трёх первых родов, процитированных Ричардом Оуэном, когда он впервые выделил группу динозавров[32].
Как бы то ни было, оба предыдущих определения выделяют динозавров как надотряд, к которому относятся два больших отряда, различающихся строением таза:
· ящеротазовые (Saurischia);
· птицетазовые (Ornithischia).
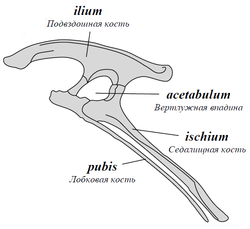
Разделение на эти два отряда было предложено в 1887 году британским палеонтологом Гарри Сили (Harry Seeley). Названия носят условный характер, так как у ящеротазовых динозавров строение таза вовсе не похоже на строение его у ящериц, а птицы происходят не от птицетазовых, а от ящеротазовых динозавров. Ящеротазовые — отряд динозавров с лобковыми костями, первично направленными вперед (как у крокодилов). У некоторых относительно поздних ящеротазовых лобковые кости становятся направленными назад (как у птиц). У птицетазовых лобковые кости направлены назад изначально.
Значение этих различий не ясно. Среди обеих групп имеются как четвероногие формы, так и двуногие, однако именно по этим особенностям определяется принадлежность динозавров к той или другой группе, независимо от их размеров, способа передвижения и питания.
| Различия в строении тазовых костей Птицетазовых и Ящеротазовых динозавров | |
| 
|
| Тазовые кости птицетазовых динозавров. Лобковые кости направлены назад | Тазовые кости ящеротазовых динозавров. Лобковые кости выступают вперёд |
Более объяснимы различия между группами в строении челюстей и зубов. У ящеротазовых динозавров зубы располагаются по краю челюсти в один ряд, который доходит до конца морды. Каждый зуб конической или долотообразной формы сидит в отдельной ячейке. У птицетазовых динозавров в передней части нижней челюсти располагалась лишённая зубов предзубная кость, часто отсутствовали передние зубы и в верхней челюсти. У многих птицетазовых был роговой клюв, как у черепах.
К ящеротазовым относятся тероподы («звероногие») и зауроподоморфные («ящероногие») динозавры. Подотряд тероподы включает в себя всех хищных ящеров, из которых наиболее крупным и известным является тираннозавр. Считается, что от мелких тероподов произошли птицы. Зауроподоморфные — в основном крупные динозавры с длинными шеями и хвостами и колоннообразными ногами. Они делятся на зауропод (брахиозавры, апатозавры, диплодоки и т. п.) и прозауропод, то есть предков зауропод).
Отряд птицетазовые включает следующие группы: стегозавры («пластинчатые ящеры»), у которых пластины расположены вдоль позвоночника; анкилозавры («окостенелые ящеры»), покрытые костяной броней по всему телу; цератопсы («рогатые ящеры»), наиболее известен трицератопс; пахицефалозавры («толстоголовые ящеры») с головами, покрытыми костяным панцирями, и орнитоподы («птиценогие»), для которых характерно передвижение преимущественно на двух ногах (бипедальность), их наиболее известный представитель — игуанодон.
Согласно традиционной классификации, стегозавров и анкилозавров объединяют в подотряд тиреофоров («щитоносных»), цератопсов и пахицелозавров в подотряд маргиноцефалов («бахромчатоголовых»), и третьим подотрядом считают орнитоподов. По новейшей классификации выделяют два подторяда: тиреофоров и цераподов, а в последний относят орнитоподов, цератопсов и пахицефалозавров в качестве инфраотрядов.
Эти определения даются для лучшего представления научных концепций о динозаврах, которые отражают современную филогенетическую систематику. Конкретность значения терминов предназначена для того, чтобы предотвратить путаницу в том, что означает термин «динозавры».
Отряд Ящеротазовые[править | править вики-текст]
 Тазовая кость тираннозавра
· †подотряд тероподы (Theropoda)
· †семейство герреразавриды (Herrerasauridae)
· †надсемейство целофизоиды (Coelophysoidae)
· †инфраотряд цератозавры (Ceratosauria)
· †семейство абелизавриды (Abelisauridae)
· †(нет ранга) тетануры (Tetanurae)
· †надсемейство Megalosauroidea (Spinosauroidea)
· †инфраотряд карнозавры (Carnosauria)
· †инфраотряд целурозавры (Coelurosauria)
· †надсемейство тираннозавроиды (Tyrannosauroidea)
· †семейство Coeluridae
· †семейство дриптозавриды (Dryptosauridae)
· †семейство процератозавриды (Proceratosauridae)
· †семейство тираннозавриды (Tyrannosauridae)
· †(нет ранга) орнитомимозавры (Ornithomimosauria)
· †(нет ранга) Maniraptora
· †(нет ранга) Oviraptoriformes
· †надсемейство теризинозавроиды (Therizinosauroidea)
·
·
· †семейство теризинозавриды (Therizinosauridae)
· †семейство алксазавриды (Alxasauridae)
· †(нет ранга) овирапторозавры (Oviraptorosauria)
· инфраотряд дейнонихозавры (Deinonychosauria)
·
·
· †семейство троодонтиды (Troodontidae)
· †семейство дромеозавриды (Dromaeosauridae)
· класс Птицы (Aves)
· †подотряд завроподоморфы (Sauropodomorpha)
· †род Текодонтозавр (Thecodontosaurus)
· †инфраотряд прозавроподы (Prosauropoda)
· †инфраотряд зауроподы (Sauropoda)
· †надсемейство Diplodocoidea
· †надсемейство Titanosauriformes
· †семейство брахиозавриды (Brachiosauridae)
· †(нет ранга) титанозавры (Titanosauria)
Тазовая кость тираннозавра
· †подотряд тероподы (Theropoda)
· †семейство герреразавриды (Herrerasauridae)
· †надсемейство целофизоиды (Coelophysoidae)
· †инфраотряд цератозавры (Ceratosauria)
· †семейство абелизавриды (Abelisauridae)
· †(нет ранга) тетануры (Tetanurae)
· †надсемейство Megalosauroidea (Spinosauroidea)
· †инфраотряд карнозавры (Carnosauria)
· †инфраотряд целурозавры (Coelurosauria)
· †надсемейство тираннозавроиды (Tyrannosauroidea)
· †семейство Coeluridae
· †семейство дриптозавриды (Dryptosauridae)
· †семейство процератозавриды (Proceratosauridae)
· †семейство тираннозавриды (Tyrannosauridae)
· †(нет ранга) орнитомимозавры (Ornithomimosauria)
· †(нет ранга) Maniraptora
· †(нет ранга) Oviraptoriformes
· †надсемейство теризинозавроиды (Therizinosauroidea)
·
·
· †семейство теризинозавриды (Therizinosauridae)
· †семейство алксазавриды (Alxasauridae)
· †(нет ранга) овирапторозавры (Oviraptorosauria)
· инфраотряд дейнонихозавры (Deinonychosauria)
·
·
· †семейство троодонтиды (Troodontidae)
· †семейство дромеозавриды (Dromaeosauridae)
· класс Птицы (Aves)
· †подотряд завроподоморфы (Sauropodomorpha)
· †род Текодонтозавр (Thecodontosaurus)
· †инфраотряд прозавроподы (Prosauropoda)
· †инфраотряд зауроподы (Sauropoda)
· †надсемейство Diplodocoidea
· †надсемейство Titanosauriformes
· †семейство брахиозавриды (Brachiosauridae)
· †(нет ранга) титанозавры (Titanosauria)
| Отряд Птицетазовые[править | править вики-текст]
 Тазовая кость эдмонтозавра
· †семейство гетеродонтозавриды (Heterodontosauridae)
· †подотряд тиреофоры (Thyreophora)
· †инфраотряд Stegosauria
· †инфраотряд анкилозавры (Ankylosauria)
· †(нет ранга) цераподы (Cerapoda)
· †подотряд Marginocephalia
· †инфраотряд пахицефалозавры (Pachycephalosauria)
· †инфраотряд цератопсы (Ceratopsia)
· †семейство пситтакозавриды (Psittacosauridae)
· †семейство протоцератопсиды (Protoceratopsidae)
· †семейство цератопсиды (Ceratopsidae)
· †подотряд орнитоподы (Ornithopoda)
· †семейство гипсилофодонтиды (Hypsilophodontidae)
· †инфраотряд игуанодонты (Iguanodontia)
· †семейство Iguanodontidae
· †семейство гадрозавриды (Hadrosauridae)
Тазовая кость эдмонтозавра
· †семейство гетеродонтозавриды (Heterodontosauridae)
· †подотряд тиреофоры (Thyreophora)
· †инфраотряд Stegosauria
· †инфраотряд анкилозавры (Ankylosauria)
· †(нет ранга) цераподы (Cerapoda)
· †подотряд Marginocephalia
· †инфраотряд пахицефалозавры (Pachycephalosauria)
· †инфраотряд цератопсы (Ceratopsia)
· †семейство пситтакозавриды (Psittacosauridae)
· †семейство протоцератопсиды (Protoceratopsidae)
· †семейство цератопсиды (Ceratopsidae)
· †подотряд орнитоподы (Ornithopoda)
· †семейство гипсилофодонтиды (Hypsilophodontidae)
· †инфраотряд игуанодонты (Iguanodontia)
· †семейство Iguanodontidae
· †семейство гадрозавриды (Hadrosauridae)
|
Существует большой консенсус среди палеонтологов, что ближайшими живущими родственниками динозавров являются птицы, происходящие от мелких хищных динозавров[33]. Современные исследования окаменелостей показывают, что птицы произошли от группы динозавров-тероподов во время юрского периода, и большинство палеонтологов считают их единственной группой динозавров, которая сохранилась и по сей день[34]. В связи с этим в кладистике птиц включают в число динозавров (иначе динозавры оказываются недопустимой, парафилетической группой). С точки зрения кладистики птицы — представители группы динозавров, то есть динозавры не вымерли[34].
Дата добавления: 2021-06-02; просмотров: 86; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!