Так, а кто из рекордсменов у нас есть в ранге морских Динозавров ? 6 страница
Именно на его изучении и основывались все первые гипотезы и теории о происхождении и родственных связях птиц: теория древесного («с деревьев вниз», Marsh, 1877)[22] и теория бегательного («с земли вверх», Williston, 1879)[23] возникновения полёта у птиц. Соответственно этим представлениям трактовалось и происхождение самих птиц — от триасовых текодонтов (архозавроморф) при древесной теории или от юрских бегающих тероподных динозавров при наземной теории[24].
В настоящее время археоптерикса уже не рассматривают в качестве общего предка всех современных птиц. Однако он, вероятно, имеет близкое родство с их настоящим предком[25]. Точное положение археоптерикса в эволюционном дереве определить трудно. Согласно кладистическому анализу китайских палеонтологов Xu et al. 2011 года, археоптерикс может представлять собой (вместе с видом Xiaotingia zhengi) параллельную тупиковую ветвь на общем стволе динозавров[26]. Однако более тщательный филогенетический анализ не подтвердил размещение археоптерикса среди дейнонихозавров, и поэтому его продолжают считать древнейшей и наиболее примитивной птицей (в составе группы Avialae)[27]. Тем не менее, были обнаружены более древние ископаемые останки, которые также могут быть отнесены к кладе Avialae, хотя на данный момент они рассматриваются как динозавры: Anchiornis, Xiaotingia и Aurornis[28].
Динозавры и птицы[править | править вики-текст]
См. также: Оперённые динозавры и Манирапторы

Голотип Microraptor gui с перьями
Гипотеза о происхождении птиц от динозавров впервые была выдвинута в 1868 году Томасом Гексли. Она основывалась на сравнении строения археоптерикса и маленького хищного динозавра рода Compsognathus.
С течением времени, и особенно в последние десятилетия, в связи с открытием многих видов древних птиц и оперённых динозавров-теропод большинство исследователей[24] стали склоняться к теории, согласно которой птицы произошли от динозавров-тероподов из группы манирапторов[29].

Реконструкция летающего хищного динозавра чанъюйраптора, покрытого перьями
По мере обнаружения всё большего количество ископаемых останков нелетающих тероподов, так или иначе имеющих родство с птицами, чёткая граница между «птицами» и «не-птицами» становится размытой, и провести различие между данными группами всё сложнее[30]. Если ранее одним из определяющих признаков птиц считалось наличие перьевого покрова, то ряд открытий конца XX — начала XXI веков в провинции Ляонин на северо-востоке Китая показывает, что многие мелкие тероподы, например синозавроптерикс[31] и синорнитозавр[32], имели перья[33]. Эволюционное происхождение перьев можно проследить начиная с карнотавра, покрытого многочисленными бугорками[34], похожими на фолликулы. Следующий этап виден у синозавроптерикса и Dilong paradoxus, покрытых волокнистым пухом[34]. Наконец, у каудиптерикса, протархеоптерикса, синорнитозавра, микрораптора и Changyuraptor yangi уже имелись настоящие перья[34].
| ||||||||||||||||||||||||||||||
| Кладограмма филогении птиц согласно Godefroit et al., 2013[35] |
Птиц роднит с динозаврами множество общих черт в строении скелета[12]. В последние годы среди палеонтологов достигнут консенсус, что ближайшими родственниками птиц являются дейнонихозавры — инфраотряд, включающий в себя семейства дромеозавров и троодонтид. Эти три группы были объединены в единую, получившую название Paraves.
Центральное место среди дромеозавров занимает «четырёхкрылый» микрораптор, который был способен летать либо планировать. Тот факт, что дейнонихозавры были в большинстве своём маленького размера, дал основание предполагать, что предок птиц вёл древесный образ жизни и был способен к планированию[36][37].
Среди возможных ближайших родственников птиц также рассматривается авимим — мелкий хищный динозавр из мелового периода[38].
Альтернативные версии происхождения[править | править вики-текст]

Реконструкция Longisquama insignis
Наряду с теорией происхождения от динозавров существует гипотеза, согласно которой птицы могли эволюционировать от архозавроморфов[21]независимо от линии развития динозавров, в поздней перми или раннем триасе[39]. Раньше в качестве такого предка рассматривались мелкие и более примитивные псевдозухии из группы текодонтов, теперь возможными предками считают преимущественно орнитозухид[21]. Впервые данная теория была высказана в 1913—1914 годах Робертом Брумом и Фридрихом Хюне, позднее защищалась Герхардом Хейельманом, и оставалась наиболее популярной вплоть до конца двадцатого столетия. Тем не менее, некоторые учёные, среди которых американские палеонтологи Ларри Мартин (англ.)русск. и Алан Федучия (англ.)русск., продолжают настаивать, что птицы произошли от более ранних архозавров типа лонгисквамы[39]. Они утверждают, что сходство между птицами и манирапторами является результатом конвергентной эволюции. В конце 1990-х годов Федучия и Мартин выдвинули модифицированную версию своей гипотезы, согласно которой манирапторы являются нелетающими потомками ранних птиц[40]. Данная гипотеза оспаривается большинством палеонтологов[41]. В июле 2014 года в продолжение своей гипотезы Алан Федучия в соавторстве опубликовал исследование, согласно которому Scansoriopteryx рассматривался ими как древняя птица — потомок архозавров. По мнению авторов публикации, наличие у него архаичных черт и анатомических особенностей, характерных для птиц, и перьев на конечностях свидетельствуют о возникновении полёта «сверху вниз», при котором архозавры обзавелись перьями. Также ими утверждается, что Scansoriopteryx и все птицы произошли от архозавров, таких как склеромохлус[42]. Филогенетические исследования никогда не подтверждали данную альтернативную интерпретацию, и прежние аргументы авторов публикации ранее уже неоднократно были подвергнуты критике как ненаучные[41].
Ещё одну версию происхождения птиц отстаивал российский палеонтолог Евгений Курочкин. По его мнению, предком настоящих птиц мог быть протоавис, который на 75 млн лет древнее археоптерикса[21][43]. Критики данной теории утверждают, что останки протоависа плохо сохранились и могут быть химерой[44].
В 1977 году палеонтолог Алик Уокер после изучения скелета триасового южноафриканского текодонта-крокодиломорфа сфенозуха предложил гипотезу о происхождении птиц от ранних, позднетриасовых крокодилов. Она базировалась на большом количестве общих черт строения черепа ранних крокодилов и птиц[45]. Аргументы Уокера неоднократно критиковались, и сам автор впоследствии признал гипотезу несостоятельной.
Помимо описанных выше основных теорий происхождения птиц, были выдвинуты и многие другие. Однако все эти гипотезы не принимаются сегодня всерьёз. К таковым относятся теория Фогта о родстве птиц и ящериц (1879), теория о связи птиц с птерозаврами (Оуэн, 1875; Сили, 1881), теория о происхождении птиц от эозухий (Г. П. Дементьев, 1940)[46], теория о родстве птиц и птицетазовых динозавров (Galton, 1970)[47].
Ранняя эволюция птиц[править | править вики-текст]
| Филогения по Lefèvre et al., 2014 [48] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|

Конфуциусорнис — примитивная ископаемая птица из раннего мелового периода. Китай
Наиболее ранние представители клады Avialae — птицеподобные динозавры Anchiornis huxleyi , Xiaotingia zhengi , Aurornis xui, известные по окаменелостям из геологической формации Тяоцзишань в Китае, датируемой концом юрского периода (около 160 млн лет назад)[35]. Многие из этих ранних форм обладали необычными анатомическими особенностями, которые в дальнейшем были утеряны птицами в ходе эволюции[35].
Диверсификация, или эволюционное разделение на множество разнообразных форм, началась в меловом периоде[49]. Многие из этих форм, в том числе и предки современных веерохвостых птиц, на первоначальном этапе сохраняли примитивные признаки от динозавров — крылья с когтями и зубы, которые впоследствии были редуцированы. Джехолорнис и другие древнейшие птицы обладали длинным хвостовым отделом позвоночника[49]. У представителей других, более развитых групп, например клады пигостилевых, 4—6 задних хвостовых позвонков были слиты воедино, образовав пигостиль.
Первой широко распространённой и разнообразной группой короткохвостых птиц стали энанциорнисовые, или «противоположные птицы», названные так потому, что строение их плечевых костей было противоположно строению таковых у современных птиц[50]. Примитивные энанциорнисовые не связаны общим происхождением с более прогрессивными веерохвостыми птицами и представляют отдельную ветвь эволюции. Среди внешне похожих на современных птиц и развивавшихся вместе с ними в меловом периоде энанциорнисов были представители, занявшие различные экологические ниши: от обитателей песчаных морских побережий и охотников за рыбой до ведущих древесный образ жизни и питающихся семенами[49]. Более развитые ихтиорнисы, несколько напоминающие современных чаек, обитали на морских просторах и также питались рыбой[51]. Морские птицы из отряда гесперорнисообразных из мезозоя настолько приспособились к охоте за рыбой, что в ходе эволюции утратили способность к полёту и полностью перешли к водному образу жизни. Несмотря на такую узкую специализацию, они являются одними из родственников современных птиц[49].
Эволюция современных птиц[править | править вики-текст]
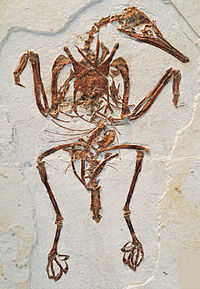
Rapaxavis pani — ископаемый вид энанциорнисовых птиц
Обнаруженные на острове Вега у берегов Западной Антарктиды останки древней птицы вегавис[52] показали, что по крайне мере одна эволюционная линия веерохвостых птиц получила развитие в начале мелового периода[52][53]и впоследствии разделилась на две основные формы, известные сегодня как подклассы бескилевых и новонёбных птиц. Бескилевые представляют относительно небольшую группу, к которой относят страусообразных и скрытохвостых. Следующей группой, разошедшейся с остальными новонёбными птицами, стали представители надотряда Galloanserae, в состав которого включают гусеобразных и курообразных. Период, когда началось данное расхождение, является предметом научных споров — большинство специалистов считают, что веерохвостые получили своё развитие в меловом периоде, а ветвь Galloanserae отделилась от остальных новонёбных до начала мел-палеогенового вымирания. Тем не менее, у этой версии имеются и противники, полагающие, что расхождение могло произойти уже после (и благодаря) гибели большей части животного мира[54]. Разногласие отчасти вызвано различными методами исследований: молекулярная генетика указывает на основную диверсификацию в середине мелового периода, а анализ окаменелостей свидетельствует о начале палеогена[54][55]. Последнее не удивительно, поскольку в первом случае речь идёт о времени отделения последнего общего предка данной группы от базальной, что произошло заведомо раньше появления останков специализированных разновидностей в палеонтологической летописи, поскольку в неё попадают только весьма распространённые в данный момент времени организмы. Более того, на момент разделения предки различных групп птиц могли быть практически неотличимы друг от друга по морфологии — всё различие между ними состояло в обусловленном генофондом эволюционном потенциале (так же, как и у различных групп примитивных плацентарных млекопитающих, которые на момент первичной радиации в палеогене были почти неотличимы друг от друга, но впоследствии дали начало совершенно различным отрядам).
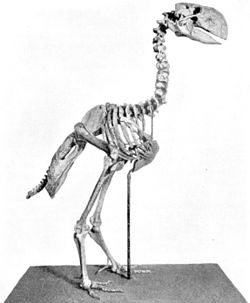
Скелет гасторниса
Древнейшей известной науке веерохвостой (настоящей) птицей является Archaeornithura meemannae, жившая около 130,7 млн лет назад на территории современного Китая[56][57].
Примерно 60 — 40 млн лет назад произошла интенсивная адаптивная радиация веерохвостых птиц, возникло большинство современных отрядов. Окаменелости птиц этого времени малочисленны и не дают представления о темпах и характере эволюции. Птицы, известные из кайнозоя, вероятно, имеют общего предка выше уровня зубастых Hesperornithiformes и Ichthyornithiformes[58]. Ископаемые новонёбные птицы современных отрядов известны преимущественно по фрагментарным отдельным остаткам из кампанских и маастрихтских отложений Монголии и Северной Америки. Это представители таких отрядов, как гусеобразные, пеликанообразные, курообразные, трубконосые, ржанкообразные, гагарообразные. Также из позднемеловых отложений США описан попугай, однако его групповая принадлежность остаётся спорной[59][60].
Бескилевые, вероятно, произошли от общего предка, отделившегося от ранних килевых форм, сохранивших палеогнатическое нёбо[61]. Форма килевой кости современных страусов предполагает, что их дальние предки летали, и что нелетающий образ жизни современных бескилевых птиц является вторичным явлением[58].
В большинстве отрядов птиц современные роды известны начиная со среднего или позднего миоцена[58]. Также многие из ныне живущих семейств птиц можно проследить по ископаемым останкам до позднего эоцена или олигоцена. Более ранние находки обычно настолько отличаются от современных родов, что установить их принадлежность к какому-либо семейству часто затруднительно[58].
Наряду с современными отрядами в кайнозое возник ряд семейств и отрядов птиц, по тем или иным причинам впоследствии вымерших[58]. Это гигантские нелетающие бескилевые эпиорнисообразные и моаобразные, а также летающие бескилевые литорнитообразные[62]. С палеоцена по плиоцен на территории Южной Америки обитали фороракосовые — вымершее семейство очень крупных хищных нелетающих птиц из отряда журавлеобразных[63]. Ещё одну группу вымерших крупных нелетающих птиц составляют Gastornithidae из палеоцена и эоцена Северной Америки, Европы и Китая. Её представители ранее выделялись в самостоятельный отряд диатримообразные, но в настоящее время рассматриваются в составе отряда гусеобразных[64].
Систематика[править | править вики-текст]
Основная статья: Систематика птиц
Первую попытку систематизации животных предпринял в IV веке до н. э. греческий учёный Аристотель — в своих сочинениях «О частях животных» и «О возникновении животных» он выделил всех известных ему птиц в «высший род» Ornithes[65][66]. Несмотря на очевидное несовершенство этой системы, вплоть до второй половины XVII века новых попыток классификации животного мира не предпринималось. В 1676 году вышла в свет рукопись «Ornithologiae libri tres», написанная английским биологом Френсисом Виллоуби, оформленная и опубликованная уже после его смерти[67]. Это первая известная работа, в которой авторы попытались классифицировать птиц на основании внешних морфологических признаков. В 1713 году была опубликована одна из первых классификаций птиц, сделанная английским биологом Джоном Рэем. Дальнейшие основы классификации птиц были заложены в 1758 году Карлом Линнеем в «Системе природы», в которой он ввёл иерархические категории и биноминальную номенклатуру при обозначении видов, используемые вплоть до настоящего времени[68]. В системе Линнея все животные были разбиты на шесть основных категорий — классов, — одну из которых заняли птицы, или Aves (позднее появились категории и более высокого порядка). В настоящее время название Aves обычно употребляется по отношению к группе, включающей в свой состав всех потомков общего предка археоптерикса и современных птиц. Альтернативная филогенетическая систематика рассматривает группу Aves в качестве клады тероподных динозавров[69]. Согласно теории происхождения птиц от динозавров, Aves и её сестринская клада Crocodilia (крокодилы) считаются единственными современными ветвями клады рептилий Archosauria[69].
Дата добавления: 2021-06-02; просмотров: 102; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!