О некоторых способах подкисленная крови 7 страница
Остановлюсь кратко и еще на двух физиологических явлениях,
напрямую связанных с ионами водорода.
Первое — об энергетике клетки. Нередко можно прочитать, что люди получают энергию непосредственно из космоса или от Солнца, что очень полезны продукты, накопившие в себе энергию нашего светила. Надо полагать, что это всего лишь красивая фантазия. Да, для поддержания жизни необходима энергия и она производится в самом организме в результате окисления кислородом жиров, белков и углеводов. От обеспечения нашего организма энергией зависит и наше здоровье, и наше долголетие. Чтобы в любом возрасте мы оставались и здоровыми, и жизнедеятельными, — мы прежде всего должны обеспечивать в полной мере свой организм энергией. Но обеспечить организм энергией вовсе не означает наполнить его жирами и углеводами
57
и, переведя математически все это в килокалории, довольствоваться достигнутым. Наш организм состоит из множества клеток и только здоровая жизнь каждой клетки может обеспечить нам полноценное здоровье. Вся совершающаяся в клетках работа — химическая, механическая, электрическая и осмотическая — выполняется с потреблением энергии. Так вот, чтобы получить необходимую для организма энергию, надо еще суметь сжечь запасенное в нем топливо. То есть надо еще доставить в организм достаточное для этого количество кислорода. Казалось бы, чего проще, ничего не надо покупать, а бери из воздуха сколько необходимо этого самого кислорода и никаких проблем. Но, оказывается, проблем здесь еще больше, чем с продуктами питания. Человек практически всю жизнь испытывает кислородное голодание (гипоксию). Я как-то слушал лекцию по этой теме (по гипоксии) и лектор сделал такое заключение, что поскольку мы ничего не можем сделать по преодолению гипоксии, то нам надо постепенно адаптировать свой организм к этому состоянию. Не было только сказано как заставить каждую клетку пользоваться меньшей долей энергии, чем ей необходимо. Но нам хорошо известно другое — при недостатке кислорода клетка может и не погибнуть, но делиться при этом она ни в коем случае не будет, а это уже прямой путь и к нашим болезням (смотрите главу 15-ю), и к преждевременному старению.
|
|
|
Почему же мы испытываем кислородное голодание? Причин для этого существует множество и познакомиться с ними можно в специальной медицинской литературе. Все эти причины я бы разделил на две группы. К первой следует отнести те, которые препятствуют насыщению крови кислородом. Самая известная из них — это понижение парциального давления кислорода во вдыхаемом воздухе. Такое может случиться не только при подъеме в горы, но в некоторых случаях и для особенно чувствительных людей и на низменных местах при резком падении барометрического давления. Но нас в данный момент интересует не эта группа причин, а другая, при которой кровь достаточно насыщена кислородом, но тем не менее отдельные органы или организм в целом испытывают кислородное голодание. Чаще всего отдельные органы испытывают такое голодание в результате атеросклероза сосудов, снабжающих их кровью. Атеросклерозу посвящена специальная глава (№10), а поэтому мы уделим сейчас внимание только кислородному голоданию всего организма, не отягченного атеросклерозом, при нормальном насыщении крови кислородом.
|
|
|
ЭФФЕКТ ВЕРИГО-БОРА
Основу разработки проблемы гипоксии заложил русский ученый-физиолог И. М. Сеченов фундаментальными работами по физиологии дыхания и газообменной функции крови. Большое значение имеют
58
также исследования русского физиолога Б. Ф. Вериго по физиологии газообмена в легких и тканях. Опираясь на идеи И. М. Сеченова о сложных формах взаимодействия между двуокисью углерода и кислородом в крови (Вериго работал в лабораториях И. М. Сеченова, И. Р. Тарханова и И. Мечникова), он впервые установил зависимость степени диссоциации оксигемоглобина1 от величины парциального давления углекислоты в крови.
|
|
|
При снижении парциального давления углекислого газа в альвеолярном воздухе и крови сродство кислорода к гемоглобину повышается, что затрудняет переход кислорода из капилляров в ткани. Это явление сегодня известно как эффект Вериго-Бора. Эффект этот был открыт независимо друг от друга Вериго (1898гр и датским физиологом Ч. Бором (1904г.)3.
Здесь я хочу ненадолго задержать внимание читателей на том, как гемоглобин крови связывает атмосферный кислород и как передает его тканям организма. При большом парциальном давлении кислорода гемоглобин (НЬ) соединяется с кислородом, образуя оксигемоглобин (НЬО2), а при низком парциальном давлении кислорода гемоглобин отдает присоединенный ранее кислород. Всю эту цепочку можно записать в виде обратимой химической реакции:
НЬ + О2 <->НЬО2
При каждом данном парциальном давлении кислорода существует определенное количественное соотношение между гемоглобином и оксигемоглобином. Если построить график зависимости количества оксигемоглобина от парциального давления кислорода, то мы получим кривую кислородной диссоциации, которая будет показывать каким образом эта реакция зависит от парциального давления кислорода. Более подробно о самом парциальном давлении говорится чуть ниже
|
|
|
в этой главе.
Но на кривую кислородной диссоциации оказывает влияние не только парциальное давление кислорода. Существенное влияние оказывает и рН крови, то есть тот самый эффект Вериго-Бора, речь о котором шла чуть выше.
' Оксигемоглобин — гемоглобин, соединенный с кислородом.
2 В это время Б. Ф. Вериго работал в Одессе в Новороссийском университете.
3 Ч. Бор — отец Нильса Бора, физика, создателя теории атома, за что ему была присуждена Нобелевская премия; Нильс Бор — отец Оге Бора, тоже физика, и тоже лауреата Нобелевской премии. Это редкий случай талантливой семьи во многих поколениях.
59
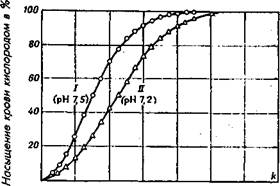
20 АО 60 80 100 !?.0 Парциальное давление кислорода в мм,рт. ст.
Рис 2.2. Кривые кислородной диссоциации для крови голубя
(по Лутцу и др., 1973.)
I — кривая, полученная при нормальных для организма птицы условиях при рН 7,5; II — кривая, полученная при всех тех же условиях, но со сдвигом рН с 7,5 до 7,2.
На рис 2.2 изображены две кривые кислородной диссоциации, которые получены для одной и той же крови и при нормальных условиях по парциальному давлению, но при разных значениях рН крови. Первое, на что я хочу обратить внимание читателей при анализе рис. 2.2 — это на то обстоятельство, что при различных значениях рН полное насыщение крови кислородом происходит при значительно меньшем парциальном давлении кислорода, чем оно реально существует на уровне моря или просто на равнинной местности.
А это означает, что нас не должна особенно волновать проблема насыщения нашей крови кислородом, по сути мы всегда имеем полное насыщение крови кислородом, если только мы не живем высоко в горах. А вот другая проблема — отдача кислорода тканям — нас должна особенно беспокоить. Очень часто наша кровь возвращается в легкие, не истратив даже 50% запасенного в ней кислорода. И в таком случае нам может помочь эффект Вериго-Бора. Например, при парциальном давлении кислорода в крови равном 40 мм. рт. ст. с рН 7,2 (по рис. 2.2) кровь может отдать 60% связанного кислорода, а та же кровь с рН 7,5 только 30%. Ясно, что для организма более благоприятна кровь с рН 7,2, чем с рН 7,5.
Физиологическое значение эффекта Вериго-Бора было отмечено многими исследователями. А упоминавшийся уже в этой главе русский ученый П. М. Альбицкий выдвинул даже гипотезу (1911г.), согласно ко-
60
торой парциальное давление углекислого газа в крови является важнейшим регулятором интенсивности окислительных процессов в тканях. Отсюда легко вытекает вывод, что при снижении в крови парциального давления углекислого газа нам следует ожидать нарушения обменных функций в организме и последующих всевозможных болезней.
Как видим, автор метода ВЛГД через полвека повторил гипотезу Альбицкого, но в то же время и предложил способ удержания углекислого газа в организме, чего не сделал Альбицкий. Конечно, самое интенсивное вымывание углекислого газа из организма происходит при глубоком дыхании. Поэтому Бутейко и решил волевым методом воспрепятствовать такому дыханию.
Многое мы делаем волевыми усилиями: и бегаем мы благодаря волевому преодолению своей лени, и физзарядкой мы занимаемся тоже благодаря волевому воздействию на самого себя, и точно так же мы обливаемся холодной водой, и точно так же мы достигаем волевыми усилиями всего и вся, чего хотим, поэтому нет ничего удивительного и в волевом управлении своим дыханием. Другое дело — многое ли нам дает такое волевое воздействие на дыхание? Возможно, все же следует найти причину самого глубокого дыхания и воздействовать на нее? Объяснение Бутейко причины глубокого дыхания нас не устраивает, так как оно бездоказательно. Как, например, связать переедание мяса или молока с глубоким дыханием? Или как леность, продолжительный сон или привычка к алкоголю приводят к глубокому дыханию? А что у детей считать причиной того же глубокого дыхания?
Вопросы эти не праздные уже потому, что если знать истинную причину глубокого дыхания, то тогда можно воздействовать на нее и в результате дыхание нормализуется. А если причина такого дыхания нам неизвестна, то тогда мы не в состоянии будем ее устранить и вынуждены будем прибегнуть к воздействию на само дыхание, что нам и предлагает Бутейко. Глубокое дыхание по его мнению является причиной многих болезней. Но мы не можем определить причину самого глубокого дыхания, а поэтому волевыми усилиями гасим глубину дыхания. Так родился метод волевой ликвидации глубокого дыхания. Ничего предосудительного в нем нет — не так быстро нам удается найти при-, чину того или иного явления.
И по-прежнему у нас нет ответа и на вопрос — в чем причина глубокого дыхания, и на вопрос — почему мы испытываем кислородное голодание при нормальном насыщении крови кислородом? Ответом на последний вопрос может служить эффект Вериго-Бора, согласно которому при снижении концентрации углекислого газа в крови возрастает сродство кислорода с гемоглобином, что затрудняет переход кислорода в ткани организма. Но такой ответ будет не совсем точным, так как сродство гемоглобина с кислородом зависит не просто
61
от концентрации углекислого газа в крови, а от концентрации ионов водорода в ней. Поэтому следует считать, что только недостаточное подкисление крови может быть причиной гипоксии всего организма при полном насыщении гемоглобина кислородом.
И если причиной гипоксии всего организма может быть относительно высокая щелочность крови, то и причиной глубокого дыхания тоже может быть испытываемое организмом кислородное голодание. Но более подробно все детали этого явления мы рассмотрим немного позже.
АТФ - УНИВЕРСАЛЬНОЕ КЛЕТОЧНОЕ ГОРЮЧЕЕ
И снова мы возвращаемся к энергетике клетки. Вспомним, что клетка — это отдельный микромир, имеющий четкие границы, внутри которых существует непрерывная химическая активность и непрерывный поток энергии. В переносе энергии от энергодающих химических реакций к процессам, идущим с потреблением энергии (которые собственно и составляют работу клетки), принимает участие АТФ (аденозинтрифосфат), выполняющий очень важную роль носителя энергии в биологических системах.
Как же образуется универсальное клеточное горючее — знаменитый АТФ?
Ответ на этот вопрос можно найти в статье Л. И. Верховского, имеющей, на мой взгляд, символическое название — "Кажется, рождается биопротоника (Химия и жизнь, 1990г., № 10). Я перескажу здесь очень кратко лишь ту часть этой статьи, где речь идет о протонах (или назовите их ионами водорода).
Известно, что наружная мембрана клеток поддерживает не только разность в концентрации отдельных веществ внутри и снаружи клеток, но также поддерживает и разность электрических потенциалов.
Предложенная лауреатом Нобелевской премии Питером Митчеллом теория образования АТФ утверждает, что при окислении жиров и углеводов ферментами дыхательной цепи через мембрану переносятся электрические заряды, а затем созданный мембраной электрохимический градиент протонов используется другим ферментом — АТФ-синтетазой, которая присоединяет к АДФ (аденозиндифосфат) неорганический фосфат:
АДФ + Фн <-» АТФ + Н2О
Эта реакция, но только со стрелкой, направленной справа налево, называется реакцией фосфорилирования, то есть реакцией переноса и присоединения еще одной фосфатной группы к аденозинди-фосфату. Аденозиндифосфат отличается от аденозинтрифосфата тем, что в нем находится две фосфатные группы, а в АТФ — три. На при-
62
соединение еще одной фосфатной группы к АДФ затрачивается энергия, которая и запасается в АТФ. Такое накопление энергии в АТФ достигается благодаря сопряжению реакции фосфорилирования с реакциями окисления. Получается, и это уже твердо установлено, что мембранный потенциал (а он возможен только при наличии достаточной концентрации ионов водорода в межклеточной жидкости, то есть при достаточном подкислении крови — прим. Н. Д.) — это связующее звено окисления и фосфорилирования.
И поэтому своеобразная гипоксия клеток может возникать и при резко выраженном разобщении процессов окисления и фосфорилирования в дыхательной цепи. Потребление клетками кислорода при этом может даже возрастать, однако значительное увеличение доли энергии, рассеиваемой в виде тепла, приводит к энергетическому 'обесцениванию клеточного дыхания. Возникает относительная недостаточность биологического окисления, при которой, несмотря на высокую интенсивность функционирования дыхательной цепи, образование АТФ не покрывает потребности в них клеток, и последние находятся по существу в состоянии гипоксии.
Приведенная выше реакция синтеза — гидролиза АТФ говорит нам не только о том, как образуется АТФ, но и как из него высвобождается в нужный момент энергия. И управление этой реакцией и влево, и вправо осуществляется с помощью протонов, которые перекачиваются протонными насосами или внутрь клетки, или наружу из нее. А эффективность работы этих насосов и энергообеспечение клеток при этом опять-таки будет зависеть от концентрации ионов водорода в крови.
ОБ ЭФФЕКТИВНОСТИ МЕТОДА ВЛГД
И вновь мы возвращаемся к задержке дыхания по методу ВЛГД. Теперь мы уже точно можем сказать, что организму нужен не сам по себе углекислый газ, а нужны ионы водорода, рождаемые углекислотой или любой другой кислотой. Но так как в организме постоянно имеется углекислота, то и подкисление крови осуществляется преимущественно ею. Это самый простой способ подкисления крови, но и самый неэффективный, так как углекислота слабо диссоциирует и не всегда она может создать должный уровень подкисления. Этот факт признает и Бутейко, когда говорит, что его методу больше подвластны острые формы болезни. И ясно почему — небольшим подкислением крови с помощью задержки дыхания удается снять остроту болезни, но не ликвидировать саму болезнь, так как для полного выздоровления не удается создать необходимого уровня подкисления с помощью углекислоты, задержанной в организме в результате неглубокого дыхания.
63
Это подтверждают и институты, проводившие проверку эффективности
метода ВЛГД.
Так нам постепенно удалось выяснить главное, что не сам по себе углекислый газ нужен организму, а только производимое им подкисление крови, а точнее, нужны только ионы водорода.
Приблизились мы и к ответу на вопрос — в чем причина глубокого дыхания?
ПРИЧИНА ГЛУБОКОГО ДЫХАНИЯ
Причиной глубокого дыхания следует считать постоянное кислородное голодание всего организма — в результате дыхательный центр выдает команду на интенсификацию дыхательных движений. Возникающая при этом гипервентиляция легких приводит к вымыванию углекислого газа из крови, вследствие чего понижается концентрация ионов водорода в крови. А снижение концентрации ионов водорода в крови повышает сродство кислорода с гемоглобином и тем самым затрудняется переход кислорода из крови в ткани.
Таким образом, круг замыкается — кислородное голодание организма приводит к гипервентиляции легких, а последняя приводит к сдвигу реакции крови в щелочную сторону, а такая реакция приводит к уменьшению высвобождения кислорода из гемоглобина и организм получает еще меньше кислорода. И в итоге продолжается глубокое
дыхание.
Но организму неведомо то, что следует только повысить кислотность крови и в результате гемоглобин высвободит больше кислорода. Нет, организм сориентирован только на забор кислорода из атмосферного воздуха и поэтому он постоянно 'держит палец на кнопке 'кислород и мы продолжаем по-прежнему глубоко дышать, испытывая при этом кислородное голодание.
И мы должны быть благодарны автору метода ВЛГД, что он предложил нам волевым усилием уменьшить глубину дыхания и таким образом бороться с гипоксией на этапе, когда нам неведома еще была причина этого явления. Но продолжать и сегодня считать, что в методе ВЛГД кардинально решены многие проблемы здравоохранения — это уже заблуждение.
ПРИЧИНА ПОВЫШЕННОЙ ЩЕЛОЧНОСТИ КРОВИ Итак, мы пришли к выводу, что глубокое дыхание является следствием кислородного голодания организма. А кислородное голодание является следствием чрезмерной щелочности крови. А что же является причиной повышенной щелочности крови? На первый взгляд кажется, что для необходимого уровня подкисления крови в ней нет достаточного количества углекислого газа.
64
Но это только так кажется. В действительности картина подкисления крови выглядит гораздо сложнее. Имеющегося в крови углекислого газа могло бы хватить для 'оптимального подкисления крови, если бы этому не препятствовала очень большая емкость буферной системы крови. Поэтому, понизив емкость буферной системы крови, мы могли бы сдвинуть реакцию крови в кислую сторону и без метода ВЛГД, более того, мы таким образом могли бы поддерживать оптимальную реакцию крови и обеспечивать себе безболезненную жизнь. Но, возможно, и буферная емкость крови тоже от чего-то зависит? Попытаемся все это выяснить.
БУФЕРНАЯ СИСТЕМА КРОВИ
Буферными называют системы (или растворы), рН которых не изменяется при прибавлении небольшого количества кислоты или щелочи. Буферные растворы содержат компоненты, диссоциирующие с образованием одноименных ионов, но отличающиеся друг от друга степенью диссоциации. В нашем случае это слабая угольная кислота и ее соль. В крови образуется карбонатная буферная система, состоящая из НзСОз и Са(НСОзЬ- Компоненты этой системы диссоциируют следующим образом:
Н2СО3 <н> Н + НСО3', Са(НС03)2 » Са2+ + 2НСО3'
Гидрокарбонат кальция — сильный электролит и поэтому диссоциация угольной кислоты (слабого электролита) будет подавлена в результате наличия в крови большого количества ионов НСОз", образующихся при диссоциации гидрокарбоната кальция. Таким образом, имеющаяся в крови угольная кислота не будет диссоциировать и не будет подкисливать кровь. Кроме того, сам гидрокарбонат кальция при диссоциации дает щелочную реакцию.
Величина рН буферного, раствора зависит не от концентрации кислоты и ее соли, а от их соотношения. Поэтому, чтобы повысить подкисление крови, необходимо изменить соотношение в компонентах буферной системы: или попытаться увеличить содержание углекислоты в крови, что и делается при задержке дыхания (но эти возможности, как уже говорилось выше, не очень велики), или же принять меры к снижению второго компонента буферной крови, то есть попытаться понизить содержание гидрокарбоната кальция в крови (это следует понимать как снижение уровня кальция в крови), что более эффективно по сравнению с задержкой дыхания сказывается на подкислении крови и вполне выполнимо.
Дата добавления: 2021-05-18; просмотров: 40; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!