Количество пища потребляемой разными червями
Тиунов А. В. «Оценка участия дождевых червей в трансформации органического вещества в дерново-подзолистой почве» 1993:
«В дерново-подзолистых почвах хвойно-широколиственных лесов Подмосковья пищевые рационы (при 15*0 подстилочных червей Lumbricus oastaneu3 и почвенно-подстилочных L.rubellus составляют соответственно 17.0-25.3 и 7.1-53.5 мг сухого веса опада/г живого веса в сутки.
Рацион собственно-почвенных червей Aporrectodea caliginosa, A.rosea и Octolasion lacteum составляет 375.9-832.8 мг почвы/г живого веса в сутки».
Б. А. Бызов. «Зоомикробные взаимоотношения в почве», 2005:
"По данным Б. Р. Стригановой [1980] дождевые черви Lumbrikus terrestis потребляют до 27 мг опада (лещина, вяз, ольха)/г сухой массы в сутки. Apporectodea rosea и Apporectodea caliginosa соответственно до 200 и 400 мг навоза или 1500 и 1000-2000 мг почвы, обогащённой органикой/г в сутки".
О. В. Чекановская «Дождевые черви и почвообразование». 1960:
«В глиняные горшки с почвой ёмкостью около 8-10 л помещались разные виды дождевых червей по 20 экземпляров и опавшие листья различных древесных пород по 30 г. Через 4 месяца было произведено взвешивание копролптов, образовавшихся при разных условиях; результаты этого опыта приведены в табл. 5.
Как можно видеть (табл. 5), на первом месте при питании разными листьями оказывается пашенный, а на последнем – малый красный червь. Отсюда ясно, насколько различной должна быть почвообразовательная деятельность на участках, заселённых разными видами червей».
|
|
|
В цитатах показан интересный факт – из естественных популяций, собственно-почвенные черви кушают намного больше как почвы, так и органических остатков. Это связано с тем, что из-за особенностей пищеварения эти черви усваивают из съеденной пищи меньше питательных веществ. Соответственно больше остаётся растениям и микроорганизмам в почве.
Скорость переработки органических остатков технологическими червями или навозными из местных популяций выше чем подстилочными, норниками или собственно-почвенными. И это делает их ценными для переработки больших объемов отходов сельскохозяйственного производства.
На дачных участках, где интерес сводится в основном к переработке небольших количеств отходов, для этих целей стоит использовать червей, полученных из местных диких популяций (в лесу, огороде или в отходах сельскохозяйственной фермы). Использование «диких» местных червей, привыкших к определённому климату и составу органических веществ, даёт положительные результаты.
На грядке, при «полевом вермикомпостировании» не так уж важна скорость переработки (если не стоит цель окультуривания почвы – улучшения, увеличения количества гумуса). Слишком большое количество биогумуса тоже плохо отражается на растениях.
|
|
|
Рыхлящая роль, перемещение растительных остатков, перемешивание почвы, место отложения капролитов червей разных эколого-трофических типов червей.
Рыхлящая роль, перемещение растительных остатков, перемешивание почвы, место отложения капролитов червей разных эколого-трофических типов червей – темы разные. Но говорить о них лучше в комплексе.
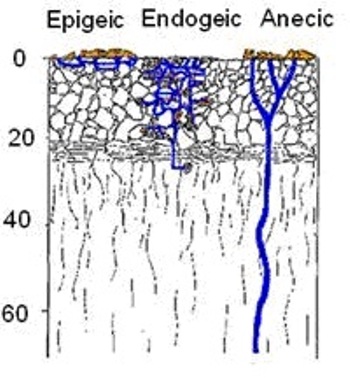
Очень наглядным для всех этих тем является рисунок 1:
Стоит обратить внимание, что на рисунке жёлтым показаны места, где черви оставляют капролиты.
Питаясь и передвигаясь в почве, дождевые черви создают систему нор, объём которых может составлять более одного процента от общего объёма верхних горизонтов почвы, а площадь поверхности – нескольких квадратных метров на 1 м2 поверхности почвы (Димо 1938; Kretzschmar 1978, 1982).
Исходя из рисунка можно сделать несколько выводов. Роющая, деятельность, перемешивание почвы с растительными остатками, затаскивание органики в почву у почвенно-подстилочных червей весьма слабая. Та же деятельность у норников сильней, чем у почвенно-подстилочных. Наиболее ярко эти виды деятельности выражены у среднеярусных червей, к которым относится обычный пашенник.
|
|
|
Та же самая информация звучит в научных работах.
А. В. Жуков «Дождевые черви как компонент биогеоценоза и их роль в зооиндикации»:
«Механическая деятельность дождевых червей, связанная с передвижением в почвенном профиле, улучшает условия воздухообмена, увлажнения, миграции питательных веществ. Так, популяция дождевого червя Aporrectodea caliginosa плотностью 100 экз./м2 способна проделать 1058 км на 1 га поверхности почвы в течение одной недели и вовлечь в механический оборот около 7,9 т грунта (Cook, Linden, 1996). Этот вид способен продуцировать в сутки около 362 мг сухого веса копролитов на 1 г сырого веса червей при температуре 5°С и до 2353 мг – при температуре 15°С».
О. В. Чекановская «Дождевые черви и почвообразование».
Исследование хорошо показывает как ведёт себя пашенник в почве – проделывает огромное количество как вертикальных, так и горизонтальных ходов и ходов под разными углами к поверхности. Много выходов на поверхность. Этим обеспечивается максимальная (по отношению к другим червям) аэрируемость почвы, перемешивание слоев. Капролиты по большей части остаются в корнеобитаемом слое почвы. А если учесть, что среднеярусные черви активно поедают органические остатки в определенной степени разложения – то они в большом количестве перемещают органику в почву – к корням растений.
|
|
|
Вот что пишет Т. Г. Добровольская «Структура бактериальных сообществ почв» 2002: «Воздействие растений на корневую биоту носит быстрый характер. Они снабжают животных корневыми легкодоступными выделениями, привлекают и отпугивают своими метаболитами. На разлагателей и инженеров экосистем растения воздействуют прямо, снабжая их органическим веществом».
Это к тому, что собственно почвенные черви привлекаются корневыми выделениями растений и оставляют капролиты (долгоиграющие удобрения) в непосредственной близости от корней. Тогда как постилочные и норники согласно рисунку 1 выносят капролиты на поверность. Их мало интересуют корневые выделения. Основной корм этих червей – слаборазложившиеся органические остатки на поверхности почвы.
«Зоомикробные взаимоотношения в почве». Б. А. Бызов. МГУ им Ломоносова.2005:
«Эпигейные (обитающие в поверхностных слоях почвы и в подстилке) черви откладывают экскременты на поверхности почвы, а обитающие в почве – в ходах или в других макропорах».
Если с подстилочными червями всё предельно ясно, то в отношении норников и собственно-почвенных червей есть определённые непонятки.
Так, Тиунов А. В. в работе «Метабиоз в почвенной системе: влияние дождевых червей на структуру и функционирование почвенной биоты» пишет:
«Экологическая роль L. terrestris до определённой степени уникальна, поскольку этот норный вид занимает промежуточное положение между собственно почвенными детритофагами и потребителями растительного опада на поверхности почвы. L. terrestris осуществляет более масштабный, по сравнению с другими почвенными беспозвоночными, вертикальный перенос вещества, и создаёт систему крупных субвертикальных каналов, соединяющих поверхность почвы и глубокие минеральные горизонты».
В той же работе читаем: «Для эндогейных червей характерно постоянное перемещение в почве, при этом вновь вырытые ходы вскоре заполняются копролитами (Kretzschmar 1983)».
Султан А. Исмаил «Вермикология», Лондон 1997:
«Дождевые черви-анецики, которые преимущественно ответственны за создание вертикальных ходов, оставляют свои копролиты, в основном, на поверхности почвы, особенно на твёрдой (Habibullah и Ismail, 1985; Krishnamoorthy, 1989), в то время как дождевые черви-эндогеики, обычно делающие горизонтальные ходы, оставляют свои копролиты внутри ходов».
То же самое мы видим на рисунке 1 и та же информация звучит в других источниках. Конечно, часть органики норники затаскивают на глубину и часть капролитов остаётся в почве. Но основная масса капролитов всё же выносится на поверхность. Тогда как основная масса капролитов собственно-почвенных червей перемещается в корнеобитаемую зону. В связи с этим не совсем понятно, почему норникам приписывают уникальные способности по вовлечению органики в почву.
Недостатком норных червей стоит считать то, что они слабо рыхлят почву, так как используют постоянные норы. Тиунов А. В. в работе «Метабиоз в почвенной системе: влияние дождевых червей на структуру и функционирование почвенной биоты» пишет: «Продолжительность существования индивидуальной норы зависит от вида червя и положения норы в почвенном профиле (Ligthart 1997).
Для эндогейных червей характерно постоянное перемещение в почве, при этом вновь вырытые ходы вскоре заполняются копролитами (Kretzschmar 1983).
Однако лабораторные и полевые наблюдения показывают, что как эндогенные, так и почвенно-подстилочные виды червей пользуются частью своих нор на протяжении длительного времени (Martin 1982; Shipitalo et al. 1988).
Норные виды, такие как L. terrestris, постоянно обитают в глубоких субвертикальных норах; по нашим наблюдениям время существования индивидуальной норы может достигать шести лет».
Как на недостаток среднеярусных червей иногда указывают на факт, что они не делают глубоких ходов. И это по видимому верно в умеренном климате. При отсутствии почвенных засух и глубокого промерзания почвы зимой среднеярусные почвы не опускаются глубоко в почву. Несколько иная картина для моего региона.
Мещерякова Е. Н. «Устойчивость дождевых червей (OLIGOCHAETA, LUMBRICIDAE, MONILIGASTRIDAE) к отрицательным температурам», 2011:
«A. caliginosa, в соответствии с холодоустойчивостью фазы червя (-5…-6°С) и коконов (-15°С), имеет обширный ареал. Напомним, что вид относится к жизненной форме собственно-почвенных (среднеярусных) червей, что предполагает их способность уходить в глубину почвы (Перель, 1979), т.е. в более мягкие температурные условия, чем вблизи поверхности (Димо, 1972). Однако в регионах с вечной мерзлотой этот вид отсутствует; нетрудно предположить, что мерзлота препятствует зимовке в глубине почвы или грунтов».
В связи с глубоким промерзанием почвы (до 3 м.) в моём регионе пашенники могут опускаться гораздо глубже пахотного горизонта, тем самым в какой-то мере дублируя деятельность норников. Глубокие вертикальные перемещения часть пашенников совершает и в засушливые периоды.
О. В. Чекановская «Дождевые черви и почвообразование». 1960:
«Мы уже говорили, что черви уходят на разную глубину в зависимости от степени засухи и промораживания почвы … В засушливых степных районах черви спускаются на значительно большую глубину».
А. В. Заушинцена, Н. В. Скалон, А. С. Заушинцен, К. С. Зубко «Реакция дождевых червей (сем. LUMBRICIDAE) на изменение абиотических факторов»:
«Иногда складываются особенно неблагоприятные условия, когда изменение влажности происходят быстро и резко, а почва может высыхать на значительную глубину. Условия длительного дефицита почвенной влаги могут привести к гибели, поэтому животные способны к продвижению за влагой в вертикальном направлении вниз почвенного профиля.
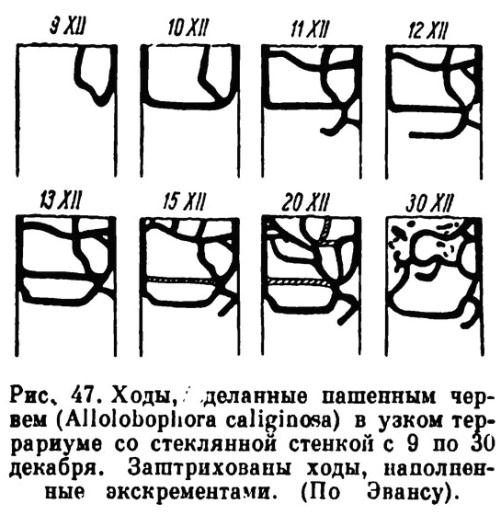
 В нашем эксперименте такой случай наблюдался в условиях сильной засухи 2012 г. При вскрытии почвенного профиля животных находили на глубине 61-100 см, в том числе и в почвообразующей породе (рис. 1). Отмечены многочисленные ходы разного диаметра. При разрушении почвенных структур обнаружены дождевые черви, как в стадии активного перемещения, так и в стадии покоя. В последнем случае они находились в основании убежища, свернувшись в клубок.
В нашем эксперименте такой случай наблюдался в условиях сильной засухи 2012 г. При вскрытии почвенного профиля животных находили на глубине 61-100 см, в том числе и в почвообразующей породе (рис. 1). Отмечены многочисленные ходы разного диаметра. При разрушении почвенных структур обнаружены дождевые черви, как в стадии активного перемещения, так и в стадии покоя. В последнем случае они находились в основании убежища, свернувшись в клубок.
… Их активная миграция, особенно вертикальная, позволяет ожидать перераспределения органического вещества и минеральных солей в почве, в результате чего в почвенном профиле будет происходить оптимизация экологических функций, их определённая направленность».
Т. С. Перель «Распространение и закономерности распределения дождевых червей фауны СССР», 1979:
«Значительно труднее разделить по характеру вертикального распределения в почве формы, питающиеся почвенным перегноем. Естественно, что все они активны в гумусовом горизонте, где содержится основной запас поступающих в почву органических веществ, в частности, в виде отмирающих корней. Известно, однако, что такие черви проникают и в нижележащие почвенные слои, способствуя углублению гумусового горизонта и, проделывая свои ходы, увеличивают скважность почвы. В известной мере это происходит и в период активной деятельности червей, но значительно глубже гумусового горизонта большинство из них уходит лишь при неблагоприятных сезонных условиях. Летом, при высыхании верхних слоев почвы, и осенью, при наступлении холодов, они мигрируют на глубину 60-90 см, где свёртываются внутри построенных из копролитов земляных капсул и впадают в состояние диапаузы».
Что касается технологических червей и навозных червей местных популяций, то для структурирования, перемешивания почвы, вовлечения в почву органических остатков они бесполезны. Их роль (так же очень важная) – переработка органических остатков на поверхности.
4. 8 Водопрочность капролитов
Чекановская «Дождевые черви и почвообразование»:
«В описанном опыте С. И. Пономарёвой оказалось, что для размывания определенного размера выбросов дождевых червей требовалось от 3.24 до 21.15 л воды, тогда как для размывания такого же размера структурных отдельностей, образованных деятельностью микроорганизмов, – от 0.005 до 1.53 л воды. Эти данные подтверждены рядом других исследователей (Бахтин и Польский, 1950; Мамытов, 1953; Соколов, 1956; Зражевский, 1957). Все они приводят очень высокие цифры водопрочности почвенных агрегатов, образованных разными видами люмбрицид в разных грунтах и при разных условиях питания. Оказалось, что из трёх видов, обычных в лесных почвах, наиболее водопрочные структурные единицы образует малый красный червь (Lumbricus rubellus), за ним идёт розовая эйзения (Eisenia rosea) и на последнем месте стоит пашенный червь (Allolobophora caliginosa)».
"Распространение и закономерности распределения дождевых червей фауны СССР" Перель Т.С.979:
"Чем выше в копролитах содержание гумуса и поглощённых оснований, тем большей водопрочностью они обладают. Этот вывод, сделанный А. А. Соколовым [1956] при сравнении выбросов; двух видов Lumbricidae» встречающихся на Северо-Западном Алтае, подтверждается и данными А. И. Зражевского, который отмечает, что копролиты у N. caliginosus и N. roseus (относящихся к собственно-почвенным дождевым червям) имеют меньшую водопрочность, чем у L. rubellus. Однако это наблюдается только в том случае, если на поверхность почвы поступает растительный опад. В вариантах опыта, в которых опад не был внесён, копролиты L. rubellus отличались даже меньшей водопрочностью, чем у’ N. caliginosus".
Стоит отметить такой факт: после многолетнего использования компостных дорожек и мульчи на грядках создалась довольно устойчивая структура. Распыления нет, после дождей или поливов структура не разрушается. Полагаю, что в моём случае не требуется максимально водостойкая структура по тому, что интенсивной обработки нет. Черви присутствуют в большом количестве и ежесекундно восстанавливают структуру.
Скорость размножения
По скорости размножения разных дождевых червей в сети есть разноречивая информация. Но в общем показано, что скорость размножения компостных, навозных червей намного выше, чем скорость размножения червей естественных популяций, особенно норников и собственно-почвенных червей.
Высокая скорость размножения бесспорно очень важна для производства удобрений и продажи червей (использования их для кормления домашней живности). Если мы говорим о «полевом вермикомпостировании», то этот показатель не так важен. При использовании одной технологии на протяжении нескольких лет, численность червей устанавливается на уровне «ёмкости среды» – то есть то оптимальное количество червей, которое может комфортно проживать в данных условиях. В этом случае количество червей ограничено количеством питания для червей, а не скоростью их размножения. Я не беру в учёт кротов, которые сокращают количество червей – в моём регионе кротов просто нет.
Дата добавления: 2019-09-08; просмотров: 189; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!