Дождевые черви, их роль в саду, огороде, поле
На разных форумах Интернета нередко можно встретить пренебрежительные отзывы о значении дождевых червей в саду, огороде, поле. Причём это пренебрежение встречается как в среде сторонников синтетических минеральных удобрений, так и в среде сторонников органических методов хозяйствования. Статьи о дождевых червях для садоводов – любителей часто воспринимаются как чисто рекламные материалы, которым не стоит доверять. С другой стороны, есть сторонники вермикультуры, превозносящие этих животных «до небес». Данный реферат написан с целью уяснить, в первую очередь для самого себя, какую реальную роль выполняют дождевые черви в растениеводстве.
Для чего простому любителю обзор научной литературы по определённой теме?
Очень многие любители считают, это совершенно не нужным. Другие, наиболее креативные, считают это даже вредным – что наука всё врёт в угоду корпоративным и иным интересам, и считают, что только они правы по любому поводу.
Всегда хочется спросить таких креативных любителей: на чем основаны твои выводы? Чаще всего ответ: «на своём личном опыте». Я не спорю, человеку наблюдательному многое может подсказать личный опыт.
Но, объясните мне, на основании чего любитель объясняет, как меняется микробиоценоз под влиянием вспашки, интродукции каких-то организмов? Понятно, что у любителя нет инструментария, чтобы отследить какие микроорганизмы есть в его почве. И уж тем более нет возможности отследить динамику изменений состава и количества микроорганизмов. В таком вопросе можно опираться только на исследования учёных, у которых такие возможности есть.
|
|
|
Или, например, любитель утверждает, что стабильный гумус создают только красные дождевые черви. На основании чего такие утверждения? На основании личного сравнения молекулярных масс гумусовых веществ, произведенных красными и бесцветными червями? Вряд ли любителю доступно такое оборудование. В таком вопросе можно опираться только на исследования учёных, у которых такие возможности есть.
При подготовке данного материала в моём распоряжении не было всей существующей информации по этой теме. Поэтому буду рад информации подтверждающей, дополняющей или опровергающей сказанное в статье (telepovoleg@mail.ru).
Дождевые черви 1 Классификация.
Наверно, для практика не особо важно, какое место в классификации животных занимают дождевые черви. Но это становится нужно, когда пытаешься поглубже разобраться с каким-либо вопросом, связанным с дождевыми червями. Например, во многих исследованиях разговор идёт не о каком-то конкретном виде дождевых червей, а обо всём семействе. Если не знать классификацию – легко запутаться.
|
|
|
КЛАСС ОЛИГОХЕТЫ или МАЛОЩЕТИНКОВЫЕ КОЛЬЧЕЦЫ (OLGOCHAETA)
Большинство олигохет – обитатели почвы: более крупные из них хорошо всем известны под названием земляных или дождевых червей. Есть, однако, и пресноводные и даже морские олигохеты.
Название «дождевые черви» обозначает не какое-то одно определённое семейство олигохет; оно применяется ко всем более или менее крупным представителям пяти различных семейств этого класса, обитающим в почве, т. е. является сборным. «Дождевые черви» – это подкласс класса олигохет.
Нас в данный момент интересует одно из семейств подкласса Дождевые черви – семейство Настоящие дождевые черви, или Люмбрициды (Lumbricidae).
Семейство Люмбрициды делится на 10 родов: Род Helodrilus, Род Allolobophora, Род Perelia, Род Dendrorilus, Род Octolasion, Род Apporectodea, Род Lumbricus, Род Eisenia, Род Eiseniela, Род Dendrobaena.
Каждый род, в свою очередь включает несколько видов. Например, пашенный червь является представителем рода Apporectodea, вид Apporectodea caliginosa; Большой выползок – представитель рода Lumbricus, вид Lumbricus terrestris; Навозный червь – представитель рода Eisenia, вид Eisenia fetida.
Описываю это всё вот для чего. В научной литературе часто оперируют не названием какого-то конкретно вида дождевых червей, в названием рода или же семейства. Например, когда речь идёт о люмбрицидах (Lumbricidae), то стоит помнить, что сказанное относится как к выползкам, так и подстилочным, навозным червям и пашенникам. Все они люмбрициды – настоящие дождевые черви.
|
|
|
Но если разговор идёт о роде люмбрикус (Lumbricus), то говорится о выползках, но никак не о пашенниках.
Кроме того, в некоторых научных работах используются только латинские названия червей, и чтобы понимать, о чём речь, стоит немного ориентироваться хотя бы в названиях родов.
В дальнейшем повествовании под словами «дождевые черви» следует понимать: «настоящие дождевые черви – люмбрициды».
Существует несколько классификаций жизненных форм дождевых червей как чисто экологического, так и морфо-экологического характера. В упрощённом виде такая классификация выглядит следующим образом:
1. Черви, живущие в глубине почвы. Питаются почвенным перегноем (детритом). Непигментированные. Этот тип можно разделить на роющих червей, живущих в глубоком горизонте почв, и норных червей, имеющих постоянные ходы, однако питающихся в гумусовом горизонте.
2. Черви, обитающие на поверхности почвы (в подстилке, под упавшими деревьями и т.п.). Питаются слабо разложившимися или практически неразложившимися растительными остатками, сильно пигментированные (красные).
|
|
|
В этом типе можно выделить подстилочных червей, которые живут в слое подстилки и ни при каких обстоятельствах (даже при пересыхании или промерзании почвы) не опускаются в землю глубже 5-10 сантиметров. К этому же типу относятся почвенно-подстилочные черви, которые проникают в почву глубже 10-20 сантиметров, но только при неблагоприятных условиях.
3. Черви, питающиеся на поверхности почвы, но вне активного периода уходящие вглубь по постоянным норам (или "норники"), сильно пигментированные (красные) Эти черви делают постоянные глубокие ходы (до 1 метра и более), которые они обычно не покидают, а при питании и спаривании высовывают на поверхность почвы только передний конец тела.
4. Черви, живущие под водой. В действительности они живут также в грунте, но затопленном. Приспособлены к существованию у условиях недостатка кислорода.
В пределах перечисленных групп дождевые черви также могут значительно различаться по образу жизни, холодо- и засухоустойчивости и т.п.
В литературе можно встретить ещё такое деление люмбрицид:
Эпигейные (живущие на поверхности) черви глубоко не зарываются в почву и живут в самых верхних слоях – в основном в пожнивных остатках, которыми они питаются.
Эндогейные (живущие в почве) черви обитают в обрабатываемом слое почвы, где они прокладывают тоннели в разных направлениях в поисках благоприятных условий и пищи, разлагая отмершие корни растений и закопанные пожнивные остатки. Эти черви могут погружаться на метровую глубину в морозную зиму.
Анекейные земляные черви – выкапывает вертикальный (более 1 метра) тоннель, служащий им убежищем с открытым отверстием на поверхности, где пожнивные остатки собираются в небольшие кучки, а впоследствии закапываются и перерабатываются. Эти особи особенно эффективны в процессе передвижения пожнивных остатков в почве.

Дождевые черви являются сапрофагами и на основании особенностей их способов питания они могут быть подразделены на две группы: детритофаги и геофагии.
Детритофаги кормятся на поверхности или близ поверхности почвы. Они питаются в обогащённом органикой почвенном горизонте, поедая в основном остатки растений и отмершие корни или экскременты млекопитающих. Таких червей называют создателями гумуса.
Черви-геофаги, обитают и питаются глубоко под поверхностью земли, поглощая большие количества почвы, богатой органикой. Их обычно называют потребителями гумуса, и они представляют собой дождевых червей эндогеиков.
Отдельные представители
Выползок обыкновенный – Lumbricus terrestris Linnaeus, 1758.

Другие названия – обыкновенный дождевой червь, большой выползок, большой красный червь. Это типичный норник. Норки этого червя можно обнаружить и подстеречь момент, когда он, возвращаясь с ночной "охоты" затаскивает туда полуразложившиеся фрагменты растений, а иногда и практически зелёные листья. Это крупный червь, достигающий 30 см в длину. Космополит.
Выползок малый – Lumbricus rubellus Hoffmeister, 1843

Другие названия – малый красный (красноватый) червь, большой подстилочный червь. Родственник большого выползка. Как явствует из названия, он существенно мельче – 5-15 см в длину. Нор этот дождевой червь не роет, а обитает в подстилке. Космополит. Иногда его называют также подлистником, путая с близким родственником – Lumbricus castaneus, который очень похож на него, но мельче – 3-8.5 см.
Большой серый червь – Aporrectodea longa (Ude, 1826)

Крупный червь (9-17 см). Космополит. В России встречается в Европейской части. Норник. Предпочитает освоенные почвы. Устаревшие названия и синонимы – Nicodrilus longus (Ude, 1826), Allolobophora longa (Ude, 1826), Allolobophora terrestris longa (Ude, 1826).
Рыжий дождевой червь – Dendrodrilus rubidus (Eisen, 1874)

Другое название – ложный навозный. Космополит. Встречается в богатых органикой верхних слоях почвы вблизи хозяйственных построек. Также как предыдущий вид заходит далеко на север. Имеются две формы: tenius, длиной 1.5-5.5 см и более крупная – subrubicundus, длиной 5-9 см.
Устаревшие названия и синонимы для D.r.tenius – Bimastus constrictus (Rosa, 1845), Bimastus tenius (Eisen, 1874), Dendrobaena rubida (Savigny, 1826), для D.r. subrubicunda – Dendrobaena subrubicunda (Eisen, 1874).
Навозный червь – Eisenia foetida Savigny, 1826

Другие названия – компостный червь, зловонный червь. Космополит. Хорошо отличается благодаря "полосатой" окраске. Длина 6-13 см. Обитает в верхних слоях почвы, богатых гумусом.
Пашенный червь Apporectodea caliginosa

Обычно является доминирующим в садах и сельскохозяйственных почвах, поймах рек. Этот вид имеет две группы. Одна группа делает горизонтальные ходы и образует небольшие холмики земли около выхода норки на поверхности. Вторая группа делает глубокие вертикальные ходы и образует более крупные холмики. Длина 60-160 мм, ширина 4-7 мм. Непигментирован.
Дождевые черви 2. Роль червей.
Многим земледельцам-практикам с давних времён было известно положительное влияние дождевых червей на урожай. Г. Н. Высоцкий (1898) заметил, что в южнорусских степях растительность лучше там, где почва содержит большое количество дождевых червей. Первые экспериментальные данные по этому вопросу были сообщены в конце 19 века (Djemil, 1896; Wollni, 1897).
Опыты, произведенные М. П. Архангельским (1929), Кравковым и Голубевой (Кравков, 1931), В. П. Ладыниным (1941), В. К. Балуевым (1950) с различными растениями и в разных условиях, подтвердили и углубили эти данные.
Особенно показательны опыты В. К. Балуева, которые были поставлены с определённым видом люмбрицид в естественной почве и в условиях, очень близких к полевым.
Опыт Балуева был осуществлён следующим образом. В глиняные неглазированные горшки ёмкостью 5 л насыпалось по 3.5 кг просеянной почвы, либо огородной, либо подзолистой супесчаной, взятой с поля, занятого пропашными культурами (опыт вёлся в этих двух вариантах). В каждом варианте было по четыре сосуда, один из которых служил контролем (без червей), а в остальных трёх на весь объём сосуда было 4, 8 и 16 червей, что соответствовало 880000-3540000 особей на 1 га. Все сосуды были закопаны в землю на поле вровень с поверхностью почвы. 15-го мая во все сосуды были посеяны зёрна яровой пшеницы сорта Лютесценс 0.62. Через 15 дней после посева в каждом из сосудов было оставлено по 9 проростков пшеницы, а остальные выдернуты. Осенью был собран урожай и учтены общий вес растений, средний вес колоса, средний вес зерна и число зёрен. Каждый вариант опыта был повторён 5 раз.
Все показатели урожайности были выше в сосудах с червями, причём выяснилось, что там где больше червей, тем урожай выше, т. е. эффект присутствия в почве червей даже при очень больших плотностях их населения, оказывается положительным.
Интересен опыт Андерсона (1940), проведённый в Калифорнии. В трёх ящиках одновременно были посеяны семена газонной травы мятлика: в одном червей вовсе не было, в другом была та же почва с червями, в третьем грунт состоял только из копролитов, собранных в той же почве. Вес надземных частей растений через одинаковый срок оказался, если принять урожай в первом ящике за 100%, – во втором 271%, в третьем 463%.
Показательны опыты А. И. Зраженского (1957), который выращивал разные породы деревьев в почве с пашенными червями и без них. Молодые деревца в вазонах с почвой, содержащей червей, росли заметно лучше, чем в почве без червей при прочих равных условиях. При этом оказалось, что влияние червей очень сильно зависит от того, какие листья даются им для корма. Опавшая листва разных деревьев по-разному действует на рост сеянцев дуба. Гниющие листья сами по себе, без червей, усиливают рост сеянцев, удобряя почву при содействии микроорганизмов (табл. 10). Но при наличии пашенных червей влияние удобрения значительно повышается, причём в весьма различной степени в зависимости от характера листьев. Можно думать, что это объясняется неодинаковой степенью пригодности листьев разных деревьев для питания дождевых червей. Выше было сказано, что черви обладают тонко развитой способностью воспринимать вкусовые ощущения и очень разборчивы в пище. Кроме того, результат деятельности червей зависит от характера микробиологических процессов в почве. Положительное влияние дождевых червей на рост сеянцев разных деревьев отчётливо выразилось уже в первый год, а через два года действие их сказалось ещё сильнее.
Влияние дождевых червей на произрастание деревьев и трав различно также в разных грунтах. Так, в опытах с саженцами сосны положительное влияние червей на рост саженцев проявилось на торфе, смешанном со сланцевой породой, значительно сильнее, чем на песке и чистом торфе. Наоборот, в некоторых опытах, например при подкладывании листьев жёлтой акации и граба на почву в сосудах с сеянцами, черви или не оказывали благоприятного действия на рост сеянцев дуба, или даже несколько тормозили рост. Этот исход опыта очень поучителен так как он свидетельствует, насколько важно всестороннее знание вопроса для управления ростом растений при проведении разного рода мелиоративных мероприятий. Отрицательный результат в этих случаях объясняется, как сказано выше, слабой активностью дождевых червей при определённых видах корма.
Эффект биогумуса и червей в процессе почвообразования хорошо виден из опыта американских учёных. На открытых разработках угля в штате Огайо (США) ежегодно образуется 4000 га отвалов. Из них 40% было облесено лиственными породами деревьев. Но разложение листового опада оказалось затруднено из-за отсутствия в почве дождевых червей. Учёные решили искусственно заселить ими участок этих площадей. Было выпущено лишь по 10 особей на каждый квадратный метр. Через пять лет растения на этом участке заметно отличались своей пышностью и здоровьем, в почве данного участка появился гумус в расчёте до 16,7 т/га.
Почвенные беспозвоночные прямо и косвенно влияют на почвенное плодородие, модифицируя режим питательных элементов в почве за счёт процессов естественной гибели, экскреции продуктов жизнедеятельности, измельчения и перемешивания органических и неорганических частиц, изменения микробиологической активности.
Прямое влияние связано с процессами естественной гибели и экскреции. Масштабы поступления в почву питательных элементов при гибели животных впечатляющие, В почвах Центральной Европы выход азота за счёт гибели достигает 24 г N/м2, что сопоставимо с ежегодной дозой минеральных азотных удобрений, вносимых на поля – 100-200 кг/га.
Дождевые черви ускоряют как процессы гумификации, так и минерализации и выветривания, увеличивают оборот питательных элементов и, как следствие, плодородие почвы и продукцию растений.
В биотическом круговороте почвенные микроорганизмы и растения закрепляют химические элементы почвы в своих клетках, а дождевые черви, и другие почвенные беспозвоночные, выводят эти элементы из органического вещества растений и микробной биомассы обратно в почву и обогащают её азотом, фосфором и калием. Органика, проходя через кишечник люмбрицид, переваривается под действием энзимов и кишечной микрофлоры, разлагается до более простых соединений, структурируется; почвенные частички обогащаются гуминовыми кислотами, кальцием, магнием, фосфорной кислотой. Выявлено, что под влиянием дождевых червей микроэлементы также становятся более доступными для растений.
Поскольку дождевые черви перерабатывают большие количества поверхностного опада и других органических материалов, они влияют на химический состав почв, а так же на транспорт питательных веществ и, таким образом, на доставку их растениям. Дождевые черви осуществляют транспорт органического материала в нижележащие почвенные слои, при этом большая часть материала переваривается, мацерируется и снова выделяется. Сами дождевые черви так же вносят в почву питательные вещества в виде азотсодержащих отходов их жизнедеятельности.
Некоторые исследователи сравнивали химический состав копролитов дождевых червей с таковым окружающей их почвой. Их результаты показали, что копролиты дождевых червей имеют более высокую общую обменную ёмкость, и, в основном, они богаче по содержанию общего органического вещества, фосфора, обменного калия и магния, а также общего обменного кальция. Дождевые черви способствуют нитрификации, поскольку они способствуют росту бактериальных популяций и аэрации почвы.
Поскольку в тканях дождевых червей содержится много белка, при разложении его так же образуется нитратный азот.
Дождевые черви усваивают не все растительные остатки, экскременты и другие органические вещества, которые они перемещают с поверхности почвы. Фактически из заглатываемого количества корма переваривается лишь небольшая её часть. Заглатываемый органический материал мацерируется, перемешивается с поглощённым неорганическим материалом, проходит через кишечник и выделяется в виде копролитов, которые имеют небольшие химические изменения.
Величина отношения C:N в копролитах дождевых червей несколько выше, чем таковая в окружающей почве, при этом количества углерода и азота, выделяемые в копролиты, иногда составляют довольно большую величину.
Жизнедеятельность червей влияет на значение рН почвы. Копролиты имеют более высокое значение рН, чем окружающая почва. Стенки ходов имеют тоже выше рН, чем окружающая почва. Экскреты червей имеют нейтральное или слабо щелочное значение рН. Быстрая трансформация мочевины в аммоний ведёт к возрастанию рН за счёт поглощения протона. Кроме того, некоторые виды экскретируют кальциевые сферулы, образующиеся в кальциевых железах и в просвете кишечника. Могут ли все эти выделения существенно изменить кислотность почвы, неизвестно, поскольку черви воздействуют только на почву, непосредственно прилегающую к их ходам. Тем не менее, считают, что некоторые черви способны оптимизировать рН в местообитаниях. Это позволяет им колонизировать кислые почвы.
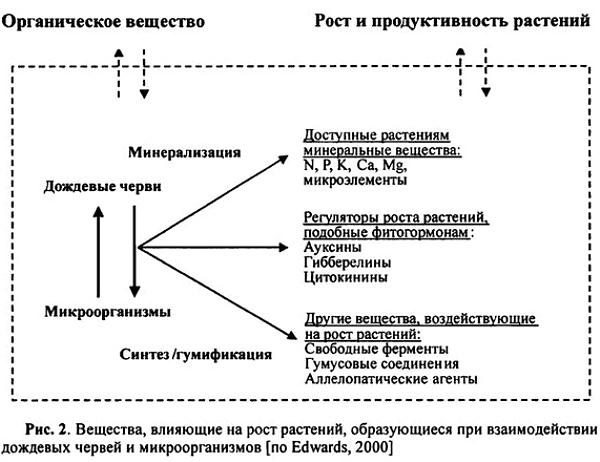
Участие червей в физических процессах, происходящих в почве, определяют следующие виды их деятельности: измельчение растительных остатков; перемещение и перемешивание подстилки и почвы; образование экскрементов; роющая деятельность.
Питаясь в разных горизонтах почвы и в подстилке, животные из группы мезо- и макрофауны активно перемешивают органические остатки с минеральными частицами. В широколиственных лесах дождевые черви – главный первичный разрушитель подстилки. Ежегодно черви перемешивают около 3-5 т подстилки/га с минеральными горизонтами, образуя так называемый гумус типа мулль. Такие же масштабы отмечены для перемешивания навоза на пастбищах. Эпигейные (обитающие в поверхностных слоях почвы и в подстилке) черви откладывают экскременты на поверхности почвы, а обитающие в почве – в ходах и в других макропорах. Годовая продукция копролитов червей в почвах умеренной зоны составляет 75-250 т/га год.
Значение того, что органическое вещество перемещается вглубь почвы червями, например, демонстрируют полевые эксперименты, проведенные в Новой Зеландии. Для утилизации навоза овец в почву интродуцировали дождевых червей. Опыты проводили с мечеными 14С-растительными остатками. Установлено, что при интродукции червей углерод аккумулировался на 10 см глубже, чем без них. Содержание углерода в почве в присутствии червей было на 30% выше. Увеличивалась продукция корней растений, растительная продукция возрастала на 70%.
Перемешивание слоев почвы идёт не только благодаря вынесению частиц почвы с глубины на поверхность; не меньшее значение имеет перемещение частиц из поверхностных слоёв в глубину. Некоторые виды дождевых червей затаскивают в свои норы огромное количество растительных остатков, что способствует образованию перегноя и возвращает в почву вещества, извлечённые из неё корнями растений.
Кроме того это может происходить путём стекания вместе с водой размоченного дождём гумусного слоя по ходам дождевых червей, а также непосредственно путем откладывания извержений в подземных ходах, подчас на большой глубине. Следует заметить, что перед свёртыванием в клубок при впадении в спячку в зимних и летних камерах обязательно опоражнивается кишечник, причем это происходит обычно неподалёку от камер, т.е. на значительной глубине.
Потребители подстилки и детритофаги при поглощении органических остатков сильно измельчают их. Дождевые черви перетирают органические остатки с помощью заглатываемых минеральных частиц в мускульном желудке. При измельчении в сотни и тысячи раз увеличивается удельная поверхность субстрата. При этом резко возрастает его доступность для микроорганизмов и, как следствие, возрастает скорость разложения.
Экскременты (копролиты) червей состоят из почвы, смешанной с органическими остатками. Они откладываются как внутри ходов червей, так и на поверхности почвы. В обоих случаях копролиты играют существенную роль в почвообразовании. Они содержат больше глинистых и илистых частиц, больше органического материала и меньше песка, чем окружающая почва, поскольку дождевые черви избирательно поглощают более мелкие частицы и органические остатки. Размер минеральных частиц, обнаруживаемых в пищеварительном тракте дождевых червей, меньше, чем в почве и составляет 100-500 мкм.
Считается, что черви не способны измельчать минеральные частицы (кроме сильно выветренных), но они перетирают растительные остатки и частицы, содержащие органоминеральные комплексы. Это – один из важнейших факторов воздействия червей на процесс разложения органического вещества.
Одно из важных воздействий дождевых червей на повышение качества почв состоит в придании ей зернистой структуры. Следует различать понятия «частицы» почвы и её «структурные отдельности» или «агрегаты». Изучение того и другого в почвах, обработанных дождевыми червями, показывает, что эти животные уменьшают размеры частиц почвы и одновременно создают почвенные агрегаты (комочки) В мышечном желудке червей происходят обкатывание и измельчение частиц почвы (особенно мягких). Механический анализ копролитов показывает, что по сравнению с исходной почвой в них содержится большее количество мелких, пылеватых частиц. Однако обнаружить их можно только после искусственного разрушения структурных отдельностей, в которые они слипаются в задних отделах кишечника червей.
Отдельные комочки извержений червей, могут сливаться друг с другом. Неоднократно ставились опыты с целью проследить, как из мелкой почвы постепенно образуются довольно крупные отдельности неправильной формы. В одном из таких опытов в деревянные ящики с почвой, просеянной через сито с ячейками в 1 мм, помещалось по семь экземпляров большого красного и пашенного червей на 1 кг почвы.
По прошествии 47 дней в почве оказалось 32.4% отдельностей размером больше 15 мм, тогда как в контрольных ящиках с почвой в тождественных условиях, но без дождевых червей их было только 3.5%. Количество частиц размером более 7 мм в почве с дождевыми червями значительно выше, а количество частиц размером менее 5 мм ниже, чем в контроле. Таким образом, структура почвы из очень мелкозернистой становится крупнозернистой и ореховатой. В дальнейшем размеры отдельностей делаются ещё несколько больше.
Отдельности в почве образуются также под влиянием деятельности других почвенных организмов, главным образом грибков и микробов; но они отличаются от образовавшихся копролитов дождевых червей более округлой формой и более рыхлой консистенцией.
Но главная особенность копролитов заключается в том их свойстве, которое почвоведы называют водопрочностью. Это название означает способность структурной отдельности почвы противостоять размыванию её водой. Водопрочность копролитов может измеряться различными методами, например количеством воды, падающей на них каплями с определённой высоты, которое необходимо для размывания их на отдельные почвенные частицы. В описанном опыте С. И. Пономарёвой оказалось, что для размывания определённого размера выбросов дождевых червей требовалось от 3.24 до 21.15 л воды, тогда как для размывания такого же размера структурных отдельностей, образованных деятельностью микроорганизмов, от 0.005 до 1.53 л воды.
Эти данные подтверждены рядом других исследователей (Бахтин и Польский, 1950; Мамытов, 1953; Соколов,167 1956; Зражевский, 1957). Все они приводят очень высокие цифры водопрочности почвенных агрегатов, образованных разными видами люмбрицид в разных грунтах и при разных условиях питания.
Следя за процессом размывания структурных отдельностей, образованных дождевыми червями, можно видеть, что в нём находятся в большом количестве неразложившиеся остатки растений (рис. 57). Очень вероятно, что чрезвычайно высокая водопрочность копролитов и их сростков объясняется наличием каркаса из структурных частей растений, связывающих их частицы. Под микроскопом видно, что отдельные части почвенной отдельности связаны спиральными волокнами. Эти спиральные волокна – растительного происхождения.
В живых стеблях трав они выстилают внутреннюю поверхность сосудов, проводящих воду и минеральные вещества от корней к листьям; они не дают спадаться их стенкам и как самые прочные части растений остаются целыми дольше других частей; после разложения стеблей они видны в изолированном состоянии. Но наличие этого органического каркаса внутри копролитов само по себе не обеспечивает ещё их водопрочности; этому содействует присутствие в них соединений кальция.
Влажные копролиты пропитаны растворами окиси кальция и его углекислой соли, которые при подсыхании кристаллизуются и цементируют мелко растёртые почвенные зернышки. А если принять во внимание, что копролит пронизан мелкими соломинками и микроскопическими волоконцами, то его исключительная водопрочность делается понятной.
Копролиты содержат в концентрированном виде ряд важных для растений веществ, как органических, так и неорганических. Водопрочность копролитов создает возможность более полного их использования растениями, так как в противном случае эти вещества быстро вымывались бы из почвы дождями. Водопрочностью обеспечивается дозированное поступление в раствор минеральных составных частей копролитов, что также выгодно для растений.
Отметим, что водопрочность не следует смешивать с водонепроницаемостью. Если бы структурные отдельности, созданные дождевыми червями, были непроницаемы для воды, наподобие нерастворимых минералов, то наличие такой структуры почвы влияло бы на плодородие почвы только отрицательно.
Исследованиями обнаружено, что удельный вес копролитов меньше удельного веса других фракций почвы. Это доказывает, что копролиты обладают большой скважностью. Вода может проникать внутрь копролита и циркулировать в нём, как по системе каналов, медленно подтачивая его и непрерывно вынося наружу небольшие порции веществ, утилизируемых микроорганизмами и высшими растениями. Капролит, по сути, работает как удобрения с пролонгированным действием.
Дождевые черви выполняют целый ряд функций, поддерживающих почвообразовательный процесс. Одна из них заключается в поддержании аэрации почвы. Необходимость интенсивного "проветривания" земляной толщи вызвана тем, что в результате жизнедеятельности биоты, а также биохимических процессов, протекающих в почве, в неё поступают различные газы.
Почва, выделяя большие объёмы различных газов, одновременно поглощает много кислорода. Без его поступления из атмосферы почвенного запаса хватило бы всего на 12-48 часов. Значит, для того чтобы предотвратить избыточное накопление одних газов и недостаточное поступление кислорода, в почве должны активно протекать процессы обмена между почвенным воздухом и атмосферой. Роль дождевых червей в улучшении газового режима состоит в том, что они проделывают в почве колоссальное количество ходов. Они в полном смысле слова пронизывают её во всех направлениях. Ходы дождевых червей очень долго не разрушаются. Их прочность создаётся за счёт уплотнения стенок и цементирования поверхности слизистыми выделениями. Поэтому они не размываются даже после очень сильных дождей. Общая длина ходов под 1 м поверхности почвы может достигать нескольких километров. Таким образом, дождевыми червями создаётся надёжный дренаж, способствующий циркуляции воздуха в почве.
Нельзя не отметить ещё одно важное следствие наличия ходов в почве. В степных засушливых районах растениям приходиться добывать влагу с больших глубин и корням, чтобы достигнуть воды, необходимо пробиваться через толщу совершенно высохшего грунта. Как установлено Г. Н. Высоцким, корни дорастают до водоносных слоёв, пользуясь ходами червей. На глубине 2 м нет ни одного корня, который бы пробил себе дорогу самостоятельно; они идут внутри ходов дождевых червей. Часто корни спаиваются вместе внутри хода, так как они оказываются сдавленными окружающим их сухим грунтом.
Ходообразование сказывается на физических свойствах почвы за счёт ряда эффектов. Черви образуют каналы между почвенными агрегатами. Ходы непрерывны в вертикальном и горизонтальном направлениях, они проникают от поверхности до глубины десятков или даже сотен сантиметров и на такое же расстояние по горизонтали. Большой диаметр ходов и их значительная протяженность делает их самыми большими порами в почве (макропорами). На их поверхности откладывается вымывающийся с поверхности почвы органический материал. В отличие от обычных пустот ходы червей изнутри покрыты соединениями, образующими мукополисахаридно-белково-липидный комплекс.
Дождевые черви за счёт роющей деятельности способствуют перераспределению порового пространства в пользу крупных пор, в которых вода меньше связана капиллярными силами. Как следствие, влажность почвы, содержащей много дождевых червей выше, чем в бедной червями почве. В почве с червями больше влаги доступной растениям. В присутствии дождевых червей возрастает инфильтрационная способность почв. Скорость инфильтрации воды в необрабатываемой почве, в которой много червей, в 6 раз выше, чем в обрабатываемой. На пастбищах, где встречается до 800 ходов червей/м2, в один ход можно залить до 100 л воды, и он не будет при этом переполнен.
Эта деятельность червей обеспечивает одновременно и полезную влажность почвы и отвод лишней влаги – сохраняет аэрацию почвы.
Стоит отметить, что в агроценозах без использования рыхления или мульчи дождевые черви с интенсивной роющей деятельностью могут способствовать некоторому высыханию почвы.
Кроме того, при изучении копролитов установлено, что они отличаются от аналогичных характеристик окружающей почвы в сторону увеличения водоудерживающей способности. Повышение влагоёмкости при копролитизации достигает 20 весовых процентов.
Рост водоудерживающей способности обеспечивается увеличением удельной поверхности копролитов, вовлечением в структуру копролитов гигроскопичного набухающего материала растительных остатков (детрита) размером 100-1000 мкм, а также капиллярным (объёмным) механизмом водоудерживания специфической агрегатной структуры копролитов.
Установлено так же, наличие червей может выступать как термостабилизирующий фактор. Hoogerkamp с сотрудниками (1983) на основе аэрофотографий, сделанных в инфракрасной области света, показал, что заселённые червями территории ночью теплее, а днём прохладнее, чем окружающие участки.
Черви и гумус
Одной из важных функций любрицид является создание гумуса.
«Зоомикробные взаимоотношения в почве». Б. А. Бызов. МГУ им Ломоносова.2005:
«Копролиты червей – это биологически активные образования, содержащие до 1010/г бактерий (это выше, чем в окружающей почве), ферменты и продукты ферментативной активности, непереваренные остатки растений, навоза и т.п. Важный компонент копролитов – гумусовые вещества. Гуминовые кислоты имеют много активных центров, связывающих катионы кальция, железа, калия, анионы, содержащие серу и фосфор. Реакции происходят благодаря донорно-акцепторным и координационным связям, образующимися между карбоксильными и другими функциональными группами гуминовой кислоты и ионами, находящимися в растворе. Питательные вещества сохраняются в гуминовых кислотах в формах, легко доступных растениям, не вымываются из почвы и высвобождаются, когда растения в них нуждаются. Растение выделяют органические кислоты, а также СО2 (протоны). Последние вытесняют катионы из мест связывания на молекуле гуминовой кислоты. За счёт медленного и постепенного высвобождения питательные вещества копролитов червей не «сжигают» корни растений».
Иногда можно услышать, что гумус создают только подстилочные и норные дождевые черви – те, что питаются подстилкой, а собственно-почвенные черви (пашенник) гумус не создают. Подтверждение такой точки зрения можно встретить в некоторых работах.
Т. С. Перель, Распространение и закономерности распределения дождевых червей Фауны СССР, 1979:
"Все дождевые черви – сапрофаги. Однако особенности их пищи могут достаточно сильно различаться. Одни способны потреблять слаборазложившиеся растительные остатки и даже практически неразложившиеся (это так называемые "гумусообразователи"), другие питаются почвенным перегноем ("гумусопотребители").
Но в той же работе Т. С. Перель пишет: «Все lumbricidae – сапрофаги. При этом одни из них, например Lumbricus terrestris, способны питаться слаборазложившимися растительными остатками и даже иногда зелёными частями травянистых растений, другие, как Nicodrilus caliginosus, – относятся ко «вторичным гумусообразователям» (по терминологии Дунгера [Dunger, 1964]), примененной им к другим систематическим группам беспозвоночных), так как они могут использовать в пищу лишь в значительной степени переработанный растительный материал».
Хочу обратить внимание, что Т. С. Пререль в одном случае называет собственно-почвенных червей «гумусопотребителями» (что может толковаться, как «не способными производить гумус»). Во втором случае тех же люмбрицид называет «вторичными гумусообразователи».
Тут поставлен знак равенства между органическим веществом почвы (почвенным перегноем) и гумусом. В этом случае, под словом «гумус» подразумевается не только гумусовые вещества, но и органика почвы в любом виде, например полуразложившиеся органические остатки. И с таких позиций всё совершенно верно.
Разночтения в вопросах гумусообразования чаще всего возникают из-за того, что однозначного определения гумусу на сегодня в науке нет. Некоторые авторы под словом «гумус» подразумевают специфические гумусовые вещества почвы. Другие же трактуют его более широко вплоть до приравнивания к детриту. Я подробно останавливался на этом вопросеhttp://sadisibiri.ru/telep-gumus.html
Стоит иметь в виду – что измельчение растительных остатков (создание детрита) и создание гуминовой и фульвокислоты, гумина – далеко не одно и то же.
В каждой конкретной цитате нужно разбираться, о чём речь идёт. Нельзя поставить знак равенства между органическими остатками разложившимися в какой-то степени и специфическим гумусовым веществом почвы.
У пашенников, как и у других собственно-почвенных червей слабей развита способность перерабатывать слаборазложившиеся органические остатки. И если под словом «гумус» иметь в виду детрит – то можно сказать, что пашенникик гумуса образует мало. Но, являясь «вторичным гумусообразователем» он перерабатывает детрит, что в итоге приводит к повышению концентрации как лабильного, так и стабильного гумуса.
Ниже привожу цитаты учёных, изучавших этот вопрос:
Тиунов А. В. «Оценка участия дождевых червей в трансформации органического вещества в дерново-подзолистой почве», 1993:
«… Проведённые исследования показали существенные различия механизмов воздействия дождевых червей сем. Luabricidae, принадлежащих к разным экологическим группам, на процесс гумификации органического вещества почвы. Если у подстилочных червей гумификация органических остатков начинается уже в кишечнике, то у собственно-почтенных червей, при питании в минеральной почве, в кишечнике происходит частичная деструкция гуминовых кислот; процесс гумификации органического материала развивается, главным образом, в экскрементах.
В кишечнике червей, благодаря высокой активности микрофлоры, наряду-с минерализацией органического вещества происходит ресинтез гумусовых соединении. Однако если подстилочные и норные черви являются типичными гумификаторами (Стриганова, 1971, 1980), то собственно-почвенные черви не могут быть однозначно отнесены к этой функциональной группе. Эти черви способны жить в бедной органическими остатками минеральной почве, извлекая энергию и пластические элементы из диспергированного в ней органического вещества, в значительной части представленного гумусовыми соединениями. Отмечены случаи, когда интродукция дождевых червой в ранее незаселённые ими агроценозы приводила к истощению почвы и падении урожайности сельскохозяйственных растения.
… Питаясь опадом, черви осуществляют его механическое разрушение и внесение в минеральные горизонты почвы. Содержание гуматов в свежих экскрементах L.rubellus достигает органического вещества – 16 и более процентов, благодаря чему в почве повышается как общее содержание органического материала, так и содержание гумусовых веществ.
В опытах с собственно-почвенными червями наблюдается иная картина. В экскрементах почвенных червей обнаружено снижение содержания гумусовых веществ по сравнению с почвой, которую заглатывали черви. Степень деструкции гумусовых соединении зависит от свойств почвы и, в частности, от содержания в ней органического вещества.
… Во всех случаях уменьшение содержания гуматов в копролитах червей происходило за счёт разрушения гуминовых кислот. Количество фульвокислот даже несколько возрастало.
Возможный механизм разрушения гумусовых веществ в кишечнике червей был описан ранее на примере тропических видов … Однако наши данные свидетельствуют, что и в умеренных широтах собственно-почвенные черви способны утилизировать стабильные гумусовые соединения, что позволяет им заселять весьма бедные органическим веществом почвы.
При питании черви активно выбирают из почвы органические компоненты. Поэтому в содержимом кишечника и в копролитах содержание органического материала всегда выше, чем в почве. Уже в кишечнике, наряду с частичным разрушением гуматов, начинается гумификация растительных остатков. Особенно активно гумификация развивается в экскрементах после их выброса в почву (Козловская, 1976; Стриганова, 1960). Благодаря этому в почве, где содержались черви, содержание гуматов постепенно повышалось, главным образом за счёт накопления в ней фульвокислот. Падение содержания гумуса в почве наблюдалось лишь при значительном увеличении (по сравнению с естественной) плотности червей в опытных сосудах (до 13.5 г ж. в./кг почвы), либо при использовании в опыте бедной почвы из A1/A2 горизонта. …
… Подстилочные черви в отличие от почвенных, не обнаруживают способности к минерализации гумусовых соединений в кишечнике. Содержание гумуса в свежих копролитах L.rubellus остается выше, чем в почве, даже при питании червей в просеянной минеральной почве. Однако в этом варианте опыта деятельность L.rubellus не приводит к увеличение содержания гумусовых веществ в почве опытных сосудов.
… Подстилочные и норные черви обеспечивают вовлечение растительного опада в минеральные почвенные горизонты и ускоряют его гумификацию. Собственно-почвенные черви принимают участие как в синтезе, так и в деструкции стабильных гумусовых веществ, составляющих основной резерв органического вещества почвы.
… В кишечнике собственно-почвенных червей, по крайней мере в случае питания в минеральных горизонтах почвы, происходит частичная минерализация гуминовых кислот. Несмотря на это, пищевая активность исследованных нами собственно-почвенных червей A. caliginosa и O.lacteum способствует накоплению в почве гумусовых соединений, что связано с развитием процесса гумификации растительных остатков в копролитах».
В приведённом отрывке стоит обратить внимание на пару моментов:
1 Собственно – почвенные черви (пашенник в том числе) создают мало гумуса если живут в почве крайне бедной органикой.
2 Норные и подстилочные, помещённые в такую почву в лабораторном эксперименте, – так же не увеличивают количество гумуса. Стоит иметь в виду, что в поле при таких условиях эти черви вообще жить не могут.
3 При достатке органических остатков собственно почвенные черви увеличивают количество гумуса.
То есть, неверно считать, что эндогейные черви (пашенник в том числе) гумус не создают.
Кроме того, в среде сторонников органических методов растениеводства нередко можно видеть мнение, что гумусированность почв напрямую приравнивается к их плодородию. Что наращивание количества гумуса – бесспорное благо, а его потеря – всегда вред.
На самом деле, как в природе, так и в агроценозах постоянно и параллельно идут процессы создания гумусовых веществ и минерализации гумуса. Одинаково нужны и те и другие процессы в определённых рамках. Для огородника основная задача получить урожай сегодня и завтра, а не только увеличить количество гумуса в почве. Нет смысла наращивать процент гумуса в почве, если это не ведёт к увеличению урожая. Получение растительной продукции – это неизбежные потери подвижного гумуса. В этом нет ничего плохого. В почве постоянно синтезируются и разрушаются гумусовые вещества, это обеспечивает жизнь экосистемы. Причем подвижный гумус для урожая более важен, чем стабильный. Кстати, образование из подвижного гумуса стабильной формы – тоже потеря подвижного гумуса для растений – он извлекается из оборота. Важно только, чтобы потери не превышали накопление. Я уже останавливался на этом вопросе. http://sadisibiri.ru/telep-gumus.html
Считать, что эндогейные черви бесполезны в плане гумусного состояния почв на основании, что они способствуют обоим процессам – неверно. Собственно-почвенные черви способствуют как стабилизации гумуса, так и его частичной минерализации. Стабильный гумус растениям не доступен и может потребляться только после частичной или полной минерализации, чему и способствуют собственно-почвенные черви.
Полагаю, в этом мудрость природы. В степных районах, где много гумуса – мало растительных остатков на поверхности. В этих районах органика запасается в гумусе. В лесной зоне органика запасается в опаде и стеблях растений. Для каждой зоны нужны свои переработчики органического вещества. Для степной зоны – почвенные. Для леса – подстилочные.
В более поздних исследованиях есть информация, что и компостные черви способны разрушать гумус.
Тихонов В. В. «Участие дождевых червей и бактерий в модификации биологических и химических свойств гумусовых веществ», 2011:
«В опытах использовались компостные черви Eisenia fétida andrei (Bouche, 1972) породы «Русский Московский гибрид», предоставленные ООО «РОСТАГРОЭКОЛОГИЯ» (пос. Пирогово), и почвенные черви Aporrectodea caliginosa (Sav.), выделенные из окультуренной дерново-подзолистой почвы.
… Микроорганизмы способны менять структуру молекул гуминовых кислот, частично их утилизируя. В модельных экспериментах и на промышленных образцах компостов нами показано уменьшение молекулярной массы и концентрации гумусовых веществ под действием червей. Присутствие червей в субстрате: низинном торфе, листве, навозе, приводило к уменьшению молекулярных масс и концентраций гумусовых веществ через 6 месяцев работы червей.
При скармливании червям препарата гуминовых кислот (ГКт), внесённого в песок, средневесовая молекулярная масса гуминовой кислоты снижалась в 1,5-2 раза: при пассаже через пищеварительный тракт червей А. caliginosa с 8,5 кДа до 5,7 кДа (рис. 7 А), а Е. fétida – с 20 кДа до 10 кДа.
Наблюдаемое уменьшение молекулярной массы гуминовых кислот, переходящих в фосфатный буфер, при пассаже можно объяснить, наряду с деструкцией молекул полимера, функцией кальциевых желез червя, которые генерируют гидрокарбонат кальция. Ионы кальция, взаимодействуя с гуминовыми веществами, снижают их растворимость.
… Таким образом, черви с одной стороны закрепляют гумусовое вещество, делая его менее водорастворимым, с другой стороны активно разрушают его».
Цитаты совершенно чётко показывают, что и компостные черви частично разрушают гумусовые вещества.
При вермикомпостировании органических остатков технологическими червями не только происходит накопление гумуса, но и развиваются предпосылки для дальнейшего разрушения гумуса.
Шаланда А. В.: «В созревшем вермикомпосте заметно увеличивается доля псевдомонад, актиномицетов, микроорганизмов, участвующих в трансформации органических и минеральных фосфатов, нитрификаторов, аэробных целлюлозоразрушающих, а также микробов, способных к деградации гумусовых веществ».
Ещё один важный момент затрагивает Т. С. Прель (Распространение и закономерности распределения дождевых червей Фауны СССР, 1979): «Значительно большее содержание гумуса в копролитах дождевых червей по сравнению с окружающей почвой отмечают многие авторы [Пономарёва, 1950; Соколов, 1956; Зражевский, 1951 и др.], причём обнаружено, что у видов, питающихся спадом, копролиты богаче гумусом и содержат больше поглощённых оснований, чем у собственно-почвенных дождевых червей. А. А. Соколов [1956] объясняет это различным соотношением минеральных почвенных частиц и органической массы в пище люмбрицид двух этих разных морфо-экологических типов.
На примере L. terrestris показано, что даже у червей одного вида количество гумуса в копролитах [Перель, Соколов, 1964] и химический состав содержащихся в них органических веществ меняется в зависимости от качества используемой ими пищи».
По сути, чем больше органического вещества в пище дождевых червей, тем больше гумуса они производят. Это касается не только люмбрицид питающихся подстилкой, но и эндогеиков – пашенников.
Бытует мнение, что пашенники едят только почву, а растительные остатки и навоз не едят. Но я много лет в своём огороде наблюдаю наличие пашенников во влажной мульче – там, где почвы собственно нет. Не имея приборов, позволяющих увидеть, чем конкретно питаются пашенники в слаборазложившихся органических остатках, по своим наблюдениям я сделать какие-то выводы не могу. Но, тем не менее, существуют свидетельства, что собственно-почвенные черви едят не одну только почву.
Б. А. Бызов. «Зоомикробные взаимоотношения в почве». МГУ им Ломоносова.2005:
«По данным Б. Р. Стригановой [1980] дождевые черви Lumbricus terrestis потребляют до 27 мг спада (лещина, вяз, ольха)/г сухой массы в сутки. Ароrrectodea roseа и Ароrrectodea caliginosa соответственно до 200 и 400 мг навоза или 1500 и 1000-2000 мг почвы, обогащенной органикой/г в сутки».
В этой цитате чётко показано, что пашенник способен потреблять навоз, если он есть. И, кроме того, обращает на себя внимание объем потребляемой пищи норника – 27 мг и пашенника – 400 мг.
В этой же монографии читаем: «Наиболее яркие примеры важности червей в процессах почвообразования на сельскохозяйственных почвах приводятся для культивируемых почв Новой Зеландии и Голландских польдеров. В Новой Зеландии сведение лесов для пастбищ снизило продуктивность экосистемы и привело к накоплению на поверхности толстого слоя экскрементов крупного рогатого скота и овец, поскольку в результате интенсивной обработки почти не осталось червей. В 1950 г. на отдельные участки были интродуцированы черви Aporrectodea caliginosa. Через несколько лет слой навоза исчез, и улучшились физические свойства. В 1966 г. то же применили на польдерах. Уже через год после интродукции увеличились стабильность агрегатов, капиллярность и количество доступной воды. Долговременный эффект выразился в увеличении содержания гумуса и азота в минеральных горизонтах. До 10 см сформировался тёмный гранулярный и гомогенный горизонт [Lee, 1985; Маceshin, 1997]».
На просторах интернета попалась цитата, подтверждающая мои наблюдения, но ни источник, ни имя автора установить не удалось. Не знаю стоит ли этой информации доверять: «О количествах потребляемой червями пищи дают представление следующие результаты. Во влажном суглинке в термостате при 15°С A. caliginosa в Австралии потребляли 200-300 мг сухой массы почвы в сутки на 1 г собственного веса. Прохождение пищи через кишечник занимало 20 час. При наличии выбора черви потребляют 10 мг сухой массы навоза в сутки на 1 г веса тела вместе с небольшим количеством листьев. При таком питании их вес увеличивается быстрее, чем при питании одними частично разложившимися листьями, так как микроорганизмы, изобилующие в навозе, либо повышают доступность пищи, либо перевариваются вместе с ней».
О. В. Чекановская «Дождевые черви и почвообразование». 1960:
«В глиняные горшки с почвой ёмкостью около 8-10 л помещались разные виды дождевых червей по 20 экземпляров и опавшие листья различных древесных пород по 30 г. Через 4 месяца было произведено взвешивание копролитов, образовавшихся при разных условиях; результаты этого опыта приведены в табл. 5.
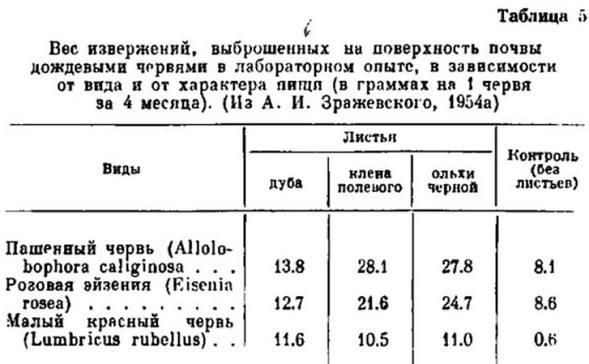
Как можно видеть (табл. 5), на первом месте при питании разными листьями оказывается пашенный, а на последнем – малый красный червь».
Эта цитата не оставляет сомнения, что пашенник способен потреблять листья деревьев, причём в большем объёме, чем подстилочный малый красный червь.
И, тем не менее, пашенник предпочитает потреблять органические остатки в определённой степени разложения, включая остатки корней.
Стригонова, «Питание почвенных сапрофагов», 1980:
«В зобе и заднем отделе кишечника А.саliginosa содержится значительное количество минеральных частиц, масса которых, однако, значительно меньше массы органики. В органическом материале преобладают компоненты, которые возможно идентифицировать, хотя они относительно мелкие и с сильно разрушенной структурой. Величина самых крупных частиц в зобе достигает 1 мм. У червей в пищевой массе преобладают остатки древесины. Кроме того в большом количестве встречаются остатки листового опада и корешков трав. Высокая степень гумификации древесных тканей в зобе позволяет предполагать, что они неоднократно подвергались ферментативной обработке в кишечнике других сапрофагов и воздействию почвенной микрофлоры. В листовых тканях имеются вкрапления аморфного гумифицированного детрита и даже минеральных частиц, которые застревают в участках с разрушенной клеточной структурой. У нескольких червей отмечены хитиновые панцири мелких членистоногих. Пищевая масса в зобе окутана слизью, мелкие органические частички у этих червей образуют скопления в каплях слизи, что наблюдалось у всех вскрытых особей. В заднем отделе кишечника возрастает масса бесструктурной органики, среди которой различаются отдельные крупные кусочки, сохраняющие структуру.
Черви, обитающие в минеральных горизонтах почвы и питающиеся детритом, не могут питаться не разрушенным листовым спадом, как представители Lumbricus. Однако они потребляют значительное количество крупных остатков растительных тканей, преимущественно корней. Высокая активность А.саliginosa в разрушении корней неоднократно отмечалась разными авторами. В наших определениях в зобе А.саliginosa найдено значительное количество остатков корней, и у некоторых особей они составляли основную массу содержимого зоба».
Т. С. Перель, Распространение и закономерности распределения дождевых червей Фауны СССР, 1979:
«Значительно труднее разделить по характеру вертикального распределения в почве формы, питающиеся почвенным перегноем. Естественно, что все они активны в гумусовом горизонте, где содержится основной запас поступающих в почву органических веществ, в частности, в виде отмирающих корней».
По сути пашенник, как и другие эндогеики, являются основными производителями гумуса из корней растений.
В среде сторонников применения вермикомпостирования чаще всего звучит утверждение, что дождевые черви играют исключительную роль в создании гумуса. Некоторые утверждают, что только дождевые черви, и никто другой, создают гумус.
В одном из источников читаем: «В природе нет других столь мощных гумусообразователей. Создать гумус другими способами пока невозможно».
Действительно, М. С. Гиляровым установлена тесная связь характера почвенного профиля, содержания и распределения в нём гумуса и мощности гумусового горизонта с видовым составом, характером миграции, биомассой почвенных сапрофагов; выявлено, что возрастание численности крупных сапрофагов и общей численности всех почвенных беспозвоночных коррелирует с повышением содержания гумуса в почве.
Но факт, что в том регионе, где я живу, в природных условиях черви живут только в переувлажнённых почвах – по берегам рек, озёр, в пониженных местах рельефа с застоем воды.
И при этом факте, у нас большую часть почв почвоведы классифицируют как черноземы – почвы, содержащие большие количества гумуса. Конечно, наши черноземы не сравняться с подобной почвой в ЦЧР. Толщина чёрного слоя, как правило, 15-20 см. Но, тем не менее, по морфологическим, физическим, химическим свойствам – это настоящий чернозём, как и повсеместно сильно истощенный антропогенным воздействием.
В общении со сторонниками исключительной роли люмбрицид в создании агрономически важного гумуса оппонентами высказывалось мнение, что в моем случае имеется гумус худшего качества – «мор» или «модер», а не мулль, который создается исключительно червями.
М. Г. Сизова «Животные как компонент почвенной экосистемы» 2006:
«С деятельностью различных групп животных почвоведы связывают образование различных форм гумуса (W. Kubiena A. Brauns и др.).
Грубый гумус-мор. Отличается кислой реакцией, малым количеством экскрементов животных. Содержит неразложившиеся и слабо разложившиеся грубые растительные остатки, пронизанные гифами грибов. Отсутствуют прочные связи органических веществ с минеральной глинистой составляющей. Такой гумус образуется во влажном и прохладном климате, когда опад богат трудно разлагаемым материалом – лигнином и целлюлозой, как под хвойными породами. В формировании такого перегноя участвуют раковинные амебы, клещи-орибатиды, ногохвостки. Участие животных незначительно, преобладает грибной тип разложения. Мор свойственен преимущественно подзолистым и дерново-подзолистым почвам.
Мягкий гумус-мулль. В его структуре заметны лишь мелкие неразложившиеся растительные фрагменты. Это результат интенсивной деятельности энхитреид, дождевых червей, многоножек, мокриц, личинок насекомых. В кишечнике животных органическое вещество растительных остатков претерпевает сложные превращения и перемешивается с минеральными частицами. В результате образуется органоминеральный комплекс. Особенно заметна роль крупных дождевых червей, которые, вовлекая растительные остатки с поверхности вглубь почвы и перемешивая их с минеральными частицами почвы, способствуют созданию зернистой структуры и углублению аккумулятивного горизонта. Кроме того, накапливаются водопрочные экскременты разных животных. Эта форма гумуса присуща серым и коричневым лесным почвам под лиственными лесами и чернозёмам.
Промежуточная форма гумуса-модер (волокнистый гумус) имеет умеренно кислую реакцию. Он образуется благодаря жизнедеятельности мелких членистоногих, экскременты которых смешиваются с минеральными частицами. В нём умеренное содержание разложившихся фрагментов растительности: структура измельчённых частей ещё различима, и они не пронизаны гифами грибов. Лучше всего эта зоогенная форма гумуса образуется в условиях повышенной влажности, которая складывается в нижних влажных слоях подстилки под более свежим опадом. В этих условиях в переработке подстилки принимают участие и крупные беспозвоночные».
То есть в чернозёмах присутствует именно мулль, иначе это не чернозём, а нечто другое. Невольно возникает вопрос: если в создании гумуса люмбрициды играют исключительную роль, то, как образовался чернозём на юге Омской области?
Этот вопрос я несколько раз задавал активным сторонникам вермикультуры. Но ответов так и не получил. Наиболее логичным объяснением было предположение одного из собеседников, что я просто не пытался найти люмбрицид в местных природных условиях. Это заставило меня разобраться в данном вопросе.
Почему на моём участке нет красных червей (выползков)?
Наука: «… Однако в конкретном биогеоценозе могут существовать только представители тех жизненных форм и в таком количестве, которые допустимы условиями этого биогеоценоза». (А. В. Жуков «Дождевые черви как компоненты биогеоценоза и их роль в зооиндикации»).
Подстилочные и роющие черви населяют места с переувлажнёнными почвами – берега водоемов, болотистые почвы, почвы влажных субтропиков. В тундре и в тайге обитают лишь подстилочные и почвенно-подстилочные формы, а в степях – только собственно почвенные.
Наука: «Наибольшей плотностью и наибольшим видовым разнообразием люмбрицид характеризуются зоны смешанных и широколиственных лесов, лесостепь и интразональные луговые ценозы. К югу от лесостепи зоомасса дождевых червей начинает снижаться. В чернозёмных почвах степей распространены только собственно почвенные формы дождевых червей и то только в почвах плакоров в очень небольшом числе и только в западных районах степной зоны. Но большая численность может быть в интразональных биотопах (в почве лугов, речных террас, на склонах оврагов) и вблизи жилья, в антропогенных биотопах (на огородах, в компостах и т. п.). («Биоиндикация почвы по беспозвоночным»).
Конкретно о червях в степной зоне Омской области читаем: «Для естественных биотопов степной зоны Омской области характерно преобладание одного-единственного вида – пашенного червя. Несмотря на то, что эта зона отличается наибольшим содержанием гумуса и величиной плодородного слоя почвы, неблагоприятный режим увлажнения и отсутствие листовой подстилки лимитируют распространение подстилочных и почвенно-подстилочных форм».
Подчёркнутое в вышеприведенном абзаце наталкивает на мысль: если создать листовую подстилку и достаточное увлажнение, то получим условия, подходящие для других червей. Но, по всей видимости, на отсутствие красных червей влияют и другие факторы, так как в берёзовых колках (где всегда есть листовой опад) червей тоже нет.
Наука: «Большая протяжённость лугово-степной зоны определяет значительную неоднородность природных условий на её территории.
Климат зоны характеризуется жарким летом и умеренно холодной зимой. В восточных областях зима более холодная и продолжительная, чем на западе. По мере продвижения на восток уменьшается количество осадков, средняя температура июля изменяется с 23-25°С на западе до 19-21°С на востоке, а средняя температура января с -4° до -25°С соответственно. Продолжительность вегетационного периода значительно больше, чем в лесостепной зоне и составляет на западе европейской части более 6 месяцев, а на востоке – около 5. В этой зоне, как и в лесостепи, часто бывают заморозки, что ещё более сокращает период вегетации растений.
Более ярко климат зоны характеризуют осадки и в целом условия увлажнения. Общее количество осадков сравнительно высокое и изменяется с запада на восток. В западных районах и Предкавказье их выпадает 500-600 мм, в Поволжье – 300-400 мм, в Западной Сибири – 300-350 мм.
На почвообразование большое влияние оказывают характер и время выпадения осадков. В основном осадки выпадают летом в виде ливней. Основная масса воды не попадает в почву, а расходуется на поверхностный сток и испарение, поэтому климат засушливый. Сухость климата усиливается постоянно дующими с юго-востока суховеями. При сравнительно небольшом количестве осадков и высокой испаряемости формируется непромывной, а в южных и восточных районах – выпотной тип водного режима».
Что собственно я и наблюдаю. Есть ряд причин, почему у нас живут только почвенные черви: засуха летом, слишком холодно зимой. Подстилочные и норники характерные для европейской части страны, просто не могут жить в таких условиях.
О. В. Чекановская Дождевые черви и почвообразование. 1960:
«Отметим, что не у всех видов люмбрицид способность переносить засуху развита в одинаковой степени. Интересны данные В. К. Балуева, который продолжал эксперименты П. Ю. Шмидта с разными видами люмбрицид. Он вёл опыты высушивания червей в почве, сравнивая разные виды.
Это давало ему возможность наблюдать реакцию их на высушивание в виде сворачивания и образования камер в почве. Оказалось, что только пашенный червь и длинная аллолобофора реагируют таким образом: 60% особей первого вида и 40% особей второго вида на третьи сутки подсушивания образовали камеры. Кроме них, лишь у особей восьмигранной дендробены в 10/6 случаев наблюдались камеры. Остальные виды при подсушивании гибли, но камер не образовывали. Таким образом, наибольшая приспособляемость оказалась у двух наиболее обычных жителей культурных земель.
… Таким образом, и по этому признаку упомянутые два вида далеко превосходят все остальные в отношении выносливости, тогда как навозный червь оказывается на последнем месте.
… На какой же именно глубине происходят зимовка и перенесение засухи дождевыми червями?
Общего ответа на этот вопрос дать невозможно. Мы уже говорили, что черви уходят на разную глубину в зависимости от степени засухи и промораживания почвы; кроме того, разные виды обладают различной способностью к пробуравливанию глубоких слоёв почвы.
В местностях с очень влажным климатом, например в Колхидской низменности (Грузия), дождевые черви, по-видимому, никогда не спускаются глубже 10-20 см. В умеренных широтах много ходов червей и их камер наблюдается на глубине 1.5-2.0 м. Сюда они спускаются, не только при неблагоприятных условиях; на этих глубинах встречаются в порядочном количестве их копролиты. В засушливых степных районах черви спускаются на значительно большую глубину.
… Вертикальные перемещения дождевых червей отнюдь не обязательно связаны с полным прекращением их активной деятельности. На значительной глубине черви могут продолжать заглатывать почву, содержащую органические вещества, и делать ходы, но деятельность их при этом становится менее интенсивной. Следовательно, между полноценной деятельностью и состоянием покоя имеются все возможные переходы».
Мещерякова Е. Н. в работе «Устойчивость дождевых червей (OLIGOCHAETA, LUMBRICIDAE, MONILIGASTRIDAE) к отрицательным температурам», 2011 делает подробный анализ влияния отрицательных температур на разных червей. В частности она пишет:
«Пороги переносимых дождевыми червями температур в процессе акклиматизации существенно меняются. Если летом все виды не выдерживают охлаждение ниже нуля, то зимовать ряд из них может при весьма низких отрицательных температурах, перенося замерзание. Таких 6 видов: A. caliginosa, D. octaedra, D. ghilarovi, E. nordenskioldi nordenskioldi, E. nordenskioldi pallida, E. sibirica. Переносимые температуры варьируют от -33°С у E. nordenskioldi nordenskioldi до -5°С у A. caliginosa (рис. 1). Не переносят замерзание и погибают при -1…-3°С 8 видов (A. rosea, D. rubidus tenuis, E. fetida, E. tetraedra L. castaneus, L. rubellus, L. terrestris, O. lacteum) …
… Норный вид L. terrestris обладает ограниченной холодоустойчивостью, но, будучи норником, зимует на глубине свыше 1.5 метров, в непромерзающих горизонтах почвы. Коконы же этот вид откладывает в камерах, расположенных в поверхностном горизонте почвы не глубже 25 см (Nuutinen, Butt, 2009). Поэтому можно предполагать, что ареал L. terrestris, в целом, ограничен низкими температурами поверхностного слоя почвы в пределах хвойно-широколиственных и широколиственных лесов».
Из этой работы становится понятным, что отсутствие норников в наших условиях обусловлено глубоким промерзанием почвы зимой. В отдельные зимы промерзание почвы достигает 2,5 метра. На моей памяти максимальное промерзание почвы составило 3 метра.
Из приведённой выше информации становится понятным, почему в природных условиях в моём регионе червей нет. Присутствуют только пашенники в агроценозах, и навозные в навозных кучах.
Остаётся открытым вопрос: если исключительная роль в создании гумуса принадлежит дождевым червям, то откуда взялся чернозём?
Стоит обратить внимание на рисунок, наглядно показывающий, кем и чем создаются разные виды гумуса.

Мулль создается только при участии мезофауны. Но мезофауна, это не только дождевые черви, но и другие животные. На рисунке они обозначены как «членистоногие» и «прочие». Как раз эти самые представители мезофауны и создают гумус там, где люмбрицид мало или их нет.
Нет повода сомневаться, что черви вносят свой вклад в гумусообразование. В литературе есть описание опытов по влиянию червей на качество гумуса.
Т. С. Перель «Распространение и закономерности распределения дождевых червей фауны СССР», 1979:
«Вегетационные сосуды (24х25 см), на дно которых был положен для дренажа слой гравия, были заполнены промытым песком, не содержащим органических веществ. На поверхность песка в каждый сосуд было внесено по 100 г (вес воздушно-сухого вещества) собранного летом опада липы и лещины (в равных соотношениях) и по 40 г опавшей хвои сосны. Позднее, в октябре, во все сосуды был добавлен собранный осенью листовой опад липы (30 г) и лещины (15 г). В середине августа в 5 сосудов было помещено по 20 экз. червей вида N. caliginosis, а затем через полтора месяца" по 3 экз. L. terrestris. Остальные 5 сосудов; были оставлены в качестве контроля. Опыт продолжался более пяти месяцев. При снятии опыта обнаружились значительные различия в потере вещества в сосудах с червями (46%) и в контрольных сосудах (31%) в результате различной скорости минерализации опада. В опыте под слоем неразложившихся листьев,, состоявшем в основном из позднее внесенного осеннего сбора опада, образовался слой однородного перегноя, в котором органические остатки потеряли свою форму. В контроле (без червей) под неразложившимися листьями находился слой перегноя типа «модер», содержащий хорошо различимые обломки листовых пластинок, черешков, хвои. В опыте песок с поверхности был хорошо гумусирован, глубже находился слабогумусированный слой с тёмно-серыми затёками. В контроле слой песка с поверхности представлял механическую смесь с обломками растительного материала, а глубже вообще не обнаруживал видимых изменений. Химические анализы показали, что участие дождевых червей в. разложении опада привело к накоплению в опытных сосудах соединений типа гуминовых кислот, содержащих значительное количество кислотных групп. В контрольном варианте произошло лишь разрушение опада и образование модероподобного слоя (Р) без заметного гумусирования песка».
Но стоит учесть, что в данном опыте не были использованы кроме червей другие почвенные животные. А они так же вносят существенный вклад с создание гумуса почв.
И. С. Белюченко, О. А. Мельник «Сельскохозяйственная экология», 2010:
«Такими же создателями гумуса, помимо дождевых червей, являются энхитреиды, а в тропических странах – термиты».
М. Гиляров «Жизнь в почве»:
«Состав почвенного населения лесов зависит от характера гумуса. Там, где гумус мягкий (мулль) и опад быстро разлагается и перемешивается, подстилка незначительна или отсутствует, а гумусовый горизонт достигает заметной мощности, отмечается обилие дождевых червей, кивсяков, мокриц, энхитреид и разнообразных личинок насекомых.
… В почвообразовании диплоподы играют большую роль. Питаются они мёртвыми растительными остатками, вовлекая в почву листовой опад, способствуют его гумификации, их экскременты становятся мелкими зернистыми структурными элементами почвы. Кроме того, в своих твёрдых покровах они накапливают много углекислого кальция, который укрепляет водопрочность почвенной структуры.
… Под микроскопом на шлифах без труда различаются экскременты основных групп микрофауны: клещей, энхитреид, нематод, ногохвосток. Работы микроморфологов показали, что в некоторых почвах практически весь гумус составляют экскременты микрофауны или продукты дальнейшего разложения этих экскрементов микроорганизмами».
Л. А. Курт, «Дети подземелья». Химия и Жизнь №4, 1991: «В кишечнике ногохвосток развивается самая разнообразная микрофлора, которая образует основу гумуса – гуминовые и фульвокислоты».
М. Г. Сизова «Животные как компанент почвенной экосистемы», 2006:
«Среди почвенных сапрофагов по их избирательности к разлагающимся органическим остаткам немецкий учёный В. Дунгер выделил первичных и вторичных разрушителей.
К первичным разрушителям он отнёс крупных почвенных и подстилочных беспозвоночных, питающихся отмершей растительностью, но не разложившейся, сохранившей тканевую структуру. Однако растительные остатки начинают перевариваться ими после выщелачивания полифенольных соединений или предварительного разрушения. Животные размельчают и мацерируют частицы растительных тканей в ротовой полости и кишечнике и разрушают связи между отдельными клетками. Они могут быстро размельчать и заглатывать мягкие ткани гниющей древесины, особенно влажной. Совершая вертикальные миграции, беспозвоночные заносят органические остатки в своих кишечниках в глубокие слои и выбрасывают их, формируя копрогенную массу. Этим они стимулируют деятельность сапротрофных микроорганизмов. Существенно то, что первичные разрушители способны переваривать структурные компоненты остатков высших растений – клетчатку, гемицеллюлозы и пектины (Б. Р. Стриганова). Но сапрофитофаги быстрее разрушают «хрупкие» виды листвы нежели остатки с сильно кутикулизированными покровами (корни, кора, листва). К активным разрушителям растительных остатков относятся красные дождевые черви, диплоподы, мокрицы, личинки длинноусых двукрылых (типулиды, бибиониды, ликорииды), некоторые виды коллембол и орибатид.
О. В. Чекановская. Дождевые черви и почвообразование. 1960:
«Однако в общем нужно сказать, что деятельность люмбрицид как образователей гумуса не имеет особого значения. Не в этом заключается их главная роль в преобразовании почв. Участие других олигохет, энхитреид, а также всех остальных почвенных животных и микробов в образовании гумуса несравненно больше. Прибавим к этому, что дождевые черви питаются остатками растений, в которых уже сильно продвинуты бактериальные процессы гниения (зелёный корм они берут крайне неохотно); поэтому их нельзя считать пионерами образования гумуса, они обрабатывают продукты, уже являющиеся, так сказать, полуфабрикатами».
Там же читаем:
«Гумификация осуществляется деятельностью не только огромного числа бактерий разных видов, но и множества мелких животных, среди которых, кроме клещей и низших насекомых, важное значение приобретают мелкие червячки (нематоды и энхитреиды), а также членистоногие (ракообразные, паукообразные, многоножки, разные отряды насекомых) Чем бы они ни питались, органические вещества, проходя через кишечник этих животных, обогащают почву.
Дождевые черви включаются в процесс почвообразования, когда гумификация почвы уже в полном разгар Мы знаем уже, что почвы, бедные гумусом, бедны и дождевыми червями; и, наоборот, чем его больше, тем обычно больше и дождевых червей.
Мы видели также, что дождевые черви хотя и участвуют в продукции гумуса, но значение их в этом отношении не так велико. В числе многих других гумификаторов они только вкладывают свою долю в продуцирование гумуса.
В известном смысле всех наземных животных можно считать гумификаторами, поскольку они обогащают почву продуктами своей жизнедеятельности, а тела их после смерти подвергаются разложению. Но их значение выступает на первый план на том этапе почвообразования, когда гумус уже создан и возникают задачи о распределении его по разным слоям почвы, о разрыхлении ее, о снабжении всей армии гумификаторов воздухом и водой, о предохранении гумуса от быстрого вымывания из почвы, об удалении избытков растительных остатков с поверхности почвы, о нейтрализации кислот, возникающих при образовании гумуса, о консервации питательных материалов для растений в водопрочных копролитах. Во всех этих процессах принимают участие и другие животные, создающие структуру и тем самым обрабатывающие почву, но главная роль в них принадлежит несомненно дождевым червям как массовым животным, мощным землероям и поглотителям почв.
Следует обратить особое внимание на то, что деятельность дождевых червей в почвообразовании органически связана с деятельностью других физических, химических и биологических агентов, участвующих в этом процессе».
«Зоомикробные взаимоотношения в почве». Б. А. Бызов. МГУ им Ломоносова.2005:
«Многие авторы отмечают относительное накопление в копролитах гумусированных химических структур, переходящих в щелочные вытяжки и придающих темную окраску последним [Курчева, 1971; Козловская, 1976]. Однако механизм гумусообразования в присутствии дождевых червей не расшифрован. Очевидно, что основные химические события в вермикомпостах, связанные с дождевыми червями, следует искать на уровне биохимической трансформации, опосредованной микроорганизмами».
В том же источнике:

Чётко показано, что образование гумуса – заслуга как животных так и микроорганизмов.
Кутовая О. В. «Влияние дождевых червей (OLIGOCHAETA, LUMBRICIDAE) на биоту и органическое вещество дерново-подзолистых почв при разных системах землепользования», 2012:
«Микроорганизмы и животное население почвы, определяющие её биологическую и биохимическую активность, обусловливают особенности формирования состава гумуса и свойств гумусовых кислот (Кононова, 1963; Курчева, 1971; Стриганова, Тиунов, 1991)».
Хомяков Н. В. «Действие пищеварительной жидкости дождевых червей на микроорганизмы», 2009:
«… Известно, что полифенолоксидазы могут участвовать как в разложении, так и в синтезе гуминовых веществ. … малый вклад самих червей, и больший – почвенных микроорганизмов, вошедших с пищей, в поддержании полифенолоксидазной активности, и, как следствие, в модификации гуминовых веществ».
А. И. Попов. «Гуминовые вещества: свойства, строение, образование»:
Гумификация – универсальное звено трансформации любых скоплений органических остатков в природе. Она может протекать в почвах, природных водах, сапропелях, торфах, при углеобразовании, образовании горючих сланцев и других каусто-биолитов.
Процесс гумификации органических остатков осуществляется при активном участии микроорганизмов, грибов и беспозвоночных животных.
Микроорганизмы и грибы способствуют превращению растительных остатков в гуминовые вещества, они продуцируют ферменты, которые катализируют процесс гумификации, к тому же эти организмы осуществляют прямой синтез гуминовоподобных соединений.
… С позиций экологии процесс гумификации является связующим звеном между совокупностью живых организмов (биоценозом) и той абиотической средой, которая стала материальной основой жизни и средой обитания. Основы экологического направления гумификации были заложены в конце 19-го столетия П. Е. Мюллером и получили дальнейшее развитие в 20-м.
П. Е. Мюллер различал два основных типа гумуса: мулль и мор, позднее для обозначения промежуточного типа Е. Раманном был введён термин «модер».
Грубый гумус – мор образуется, когда в трансформации поступающего в почву органического материала принимают участие преимущественно грибы и комплекс микроорганизмов (бактерий и протистов); средний гумус – модер образуется при участии грибов, микроорганизмов и микроскопических беспозвоночных; наконец, формирование тонкого гумуса – мулль – связано не только с грибами, микроорганизмами, микроскопическими беспозвоночными, но и с мезофауной, особенно, если последняя представлена дождевыми червями. При этом гумус типа мулль образуется из исходного органического материала при высокой интенсивности биологического круговорота биофильных элементов, а мор – при низкой.
Образование муллевого гумуса происходит также и при вермикомпостировании – переработке органического материала (чаще всего органических отходов различного происхождения) с помощью искусственного биоценотического комплекса гетеротрофных организмов, включающего в себя дождевых червей и естественно сопутствующих им представителей микроскопических беспозвоночных, а также сообщества микроорганизмов и грибов. При этом, как свидетельствует ряд авторов, данный биотехнологический приём усиливает гумификацию органического вещества по сравнению с обычным компостированием.
Усиление гумификации в присутствии почвенных беспозвоночных животных обусловлено несколькими причинами. Так, установлено, что в пищеварительной системе отдельных почвенных беспозвоночных животных (в частности, дождевых червей) наряду с минерализацией (окислением) происходит и гумификация органических остатков. В этом случае в замкнутом пространстве пищеварительной системы дождевых червей при постоянной влажности и температуре оказываются органические и минеральные вещества, активизированная микрофлора и ферменты. Таким образом, создаются оптимальные условия для новообразования ГВ. При этом ГВ, синтезированные в пищеварительной системе дождевых червей, стабилизированы в виде органоминеральных комплексов и способны долгое время сохраняться в почве.
Правда, Л. С. Козловская [156] склонна предполагать, что синтез ГВ осуществляется главным образом в копролитах дождевых червей через один-два месяца после их выброса. Обогащённые микроорганизмами копролиты, находясь в почве, становятся центрами биохимических превращений, где процессы разложения органического вещества сопровождаются синтезом ГВ.
Ещё одной причиной может быть образующийся в пищеварительной системе некоторых почвенных беспозвоночных аммиак, который способен взаимодействовать с лигнином; данное явление также имеет большое значение при гумификации. Кроме того, как было показано in vitro, пероксидазы беспозвоночных животных (земляных червей, многоножек, жуков, слизней и улиток) могут участвовать в полимеризации ароматических соединений, тем самым ускоряя гумификацию».
Данный фрагмент чётко показывает, что строго говоря, гумус черви не создают, а способствуют гумусообразованию.
Н. Ф. Ганжара «Почвоведение», 2001:
«Почвенные грибы – нитевидные одноклеточные и многоклеточные гетеротрофные, сапрофитные микроорганизмы, насчитывающие более 100 тыс. видов. В процессе питания они обладают способностью разлагать органические вещества, участвуют в минерализации растительных и животных остатков и в образовании гумусовых кислот, способных разрушать первичные и вторичные минералы в почвах».
Кутовая О. В. «Влияние дождевых червей на биоту и органическое вещество дерново-подзолистых почв при разных системах землепользования» 2012:«…практически все живые организмы почвы участвуют в трансформации органических веществ, биохимических процессах гумусообразования».
Сравнение червей разных типов
В разных источниках для садоводов-любителей, посвящённых использованию дождевых червей для растениеводства можно встретить заявления о исключительной роли отдельных видов люмбрицид. В некоторых источниках на пьедестал ставят навозных червей, в других – норников.
В большинстве случаев выводы делаются умозрительно. В редких случаях – на основе простейших опытов, не учитывающих многие переменные. И как правило, никто не опирается на исследования, сравнивающие эффекты от разных червей. Если такая информация и используется, то результаты одних исследований объявляются верными, а результаты других – заблуждением.
Ниже привожу информацию по сравнению разных червей ничего не фильтруя и не искажая. На момент работы над этим материалом мне были доступны не все источники на эту тему. Буду рад, если кто-то предоставит и другую информацию.
Влияние на микробиоценоз
А. В. Жуков. «Дождевые черви как компонент биогеоценоза и их роль в зооиндекации»:
«Эффект дождевых червей на почвенные процессы различается между экологическими группами и видами дождевых червей. Дождевые черви Lumbricus terrestris и другие норники могут рассматриваться как одни из наиболее функционально деятельных почвенных животных умеренной зоны».
Нет повода сомневаться что для умеренной зоны так оно и есть. Но, не все так однозначно для степной зоны.
Тиунов А. В. «Метабиоз в почвенной системе: влияние дождевых червей на структуру и функционирование почвенной биоты»:
«Экологическая роль L. terrestris до определённой степени уникальна, поскольку этот норный вид занимает промежуточное положение между собственно-почвенными детритофагами и потребителями растительного опада на поверхности почвы. L. terrestris осуществляет более масштабный, по сравнению с другими почвенными беспозвоночными, вертикальный перенос вещества, и создает систему крупных субвертикальных каналов, соединяющих поверхность почвы и глубокие минеральные горизонты. Однако основные механизмы влияния L. terrestris на другие компоненты почвенной биоты весьма просты и неспецифичны. Это позволяет утверждать, что выводы нашей работы в основном отражают общие закономерности влияния крупных почвенных беспозвоночных – средообразователей на структуру и функционирование почвенной биоты.
… Сравнение данных из варианта 2 нашего эксперимента с данными по микробной активности и динамике минеральных форм азота в копролитах эндогейных червей Aporrectodea caliginosa (Scheu 1987, 1987; Тиунов 1989, 1991, 1993) показывает, что эндогейные и норные черви при питании минеральной почвой оказывают на неё сходное воздействие.
… Таким образом, влияние дождевых червей разных морфо-экологических групп на микробную активность почвы (биомасса, дыхание) и на динамику минеральных форм азота по-видимому мало специфично.
… Таким образом, ключевые характеристики микробной активности в копролитах определяются, прежде всего, свойствами потребляемых червями субстратов».
Совершенно ясно показано, что основное отличие норников от всех остальных червей в том, что они проделывают глубокие норы. Но нет большой разницы между влиянием на почвенную биоту разных червей, если они питаются и в почве и в подстилке.
Романова Е. М., Игнаткин Д. С., Мухитова М. Э., Романов В. В., Шленкина Т. М. «Исследование симбиотической микробиоты представителей вида Lumbricus terrestris и оценка перспектив использования их в качестве вермикультуры для биодеструкции органических отходов сельскохозяйственного производства»:
"… Сравнительные исследования показали, что кишечный микробиоценоз L. terrestris включал в 1,5 раза больше микроорганизмов, чем ферментированный субстрат, подлежащий переработке, но значительно уступал по количеству симбионтной микробиоты промышленной линии E.f. andrei.
… Количественные исследования целлюлозолитических бактерий показали, что в содержимом кишечника L. terrestris их численность в 2,7 раза была выше, чем в ферментированном субстрате, и в два раза выше, чем у промышленной вермикультуры E.f. andrei.
… По данным литературы, в ходе вермикомпостирования субстрата он претерпевает существенные изменения. В нем не только увеличивается общая численность грибов, но и изменяется структура сообщества микромицетов в сторону увеличения доли сапрофитных грибов (Trichoderma) за счёт сокращения доли патогенных грибов [7, 8, 9, 10].
В ходе проведённых исследований нами было установлено, что численность сапрофитных грибов в микробиоценозе кишечника L. terrestris в 2,8 раза больше, чем в исходном субстрате. В кишечном микробиоценозе промышленной вермикультуры E.f. andrei численность сапрофитных грибов достоверно ниже, чем у почвенных червей.
Актиномицеты – обязательный компонент кишечного микробиоценоза дождевых червей. Благодаря актиномицетам, вырабатывающим антибиотические вещества, дождевые черви ничем не болеют [11].
По результатам наших исследований, наиболее высокая численность актиномицетов в кишечном микробиоценозе L. terrestris. Численность актиномицетов в кишечном микробиоценозе промышленной линии E.f. аndrei достоверно ниже, чем у L. terrestris. В субстрате актиномицетов в 2,6 меньше, чем в кишечном микробиоценозе L. terrestris, и в 2 раза меньше, чем у промышленной линии E.f. аndrei (рис. 4.), (р<0,05).
Актиномицеты – активные деструкторы сложных субстратов, которые они преобразуют в вещество гумуса [12].
… Органические соединения фосфора разлагаются бактериями родов Pseudomonas, Bacillus (B. megaterium, B. mesentericus), грибами, некоторыми актиномицетами, дрожжами (Candida, Saccharomyces, Rhodotorula и другие) [12].
По результатам наших исследований в кишечном микробиоценозе L. terrestris микробиоты, мобилизующей органофосфаты, достоверно меньше, чем в микробиоценозе кишечника промышленной линии E.f. andrei, однако в 1,7 раза больше, чем в субстрате, подлежащем переработке.
По данным литературных источников, фосфор находится в почве в виде нерастворимых органических и минеральных соединений, на 95% недоступных растениям [13], только благодаря эффективной работе симбионтной микробиоты, которая переводит их в растворимое состояние, фосфор становится доступным.
Биодеструкцию белков в аммиачный азот осуществляют в основном микроорганизмы, относящиеся к родам: Pseudomonas (P. florescens, P. aeruginosa), Proteus (P.vulgaris), Bacillus (B. mycoides, B. subtiis, B. cereus) и Closstridium (C. putrifius, C. sporogenes), а также актиномицеты и грибы, выделяющие протеолитические ферменты, расщепляющие белки до аминокислот, которые в клетках дезаминируются с образованием аммиака, органических кислот и других продуктов [12].
Исследование численности аммонифицирующей микробиоты показало, что в микробиоценозе L. terrestris её содержание колеблется на уровне 9-11 млн. КОЕ/г, что достоверно меньше, чем в содержимом кишечника промышленной линии E.f. andrei, но значительно выше (в 1,7 раза), чем в субстрате, подлежащем биодеструкции.
Нитрифицирующая микробиота является одной из важнейших для дождевых червей групп микроорганизмов, поскольку защищает их организм от повышенных концентраций аммиака. Нитрификаторы – это в основном бактерии семейства Nitrobactericidae, которые в два этапа осуществляют окисление аммиака до азотной кислоты [8, 12].
При оценке численности нитрифицирующей микрофлоры в кишечном микробиоценозе L. terrestris её содержание колеблется и составляет около 3 млн. КОЕ/г, что в 1,3 раза больше, чем в субстрате, подлежащем деструкции, но значительно меньше, чем в микробиоценозе промышленной линии.
Выводы. Подводя итог проведённым исследованиям, необходимо отметить, что дождевые черви L. terrestris Средневолжского региона обладают всем спектром необходимых для вермикультуры микроорганизмов, способных осуществить эффективную биотрансформацию субстрата. По ряду показателей микробиоценоз L. terrestris даже богаче микробиоценоза промышленной линии E.f. andrei. В частности, микробиоценоз почвенных червей содержит больше целлюлозолитических бактерий, сапрофитных грибов, актиномицетов, но уступает промышленной линии по количеству микробиоты, мобилизующей органофосфаты, а также микробиоты, осуществляющей нитрификацию и аммонификацию. Все полученные результаты свидетельствуют в пользу перспективности использования представителей вида L. terrestris в качестве вермикультуры для утилизации отходов животноводства, содержащих повышенные количества целлюлозы и лигнина. Для расширенного эффективного использования представителей вида L. terrestris в качестве вермикультуры для утилизации всех видов отходов животноводства недостаточно микробиоты, ответственной за процесс быстрой биотрансформации органических и минеральных форм азота и фосфатов».
Прусак А. В., Смагин А. В., Костина Н. В., Умаров М. М., Богданова Т. В. «Гидрофизические свойства и биологическая активность капролитов дождевых червей разных эколого-трофических групп», 2008:
«В лабораторных условиях нами проводился модельный эксперимент по инкубации дерново-подзолистой легкосуглинистой почвы с целью дифференцированной оценки влияния культур дождевых червей разных эколого-трофических групп: почвенно-подстилочных (Lumbricus rubellus) и типично почвенных (Aporrectodea rosea) на гидрофизические свойства и биологическую активность почвы.
… Оценка функциональных показателей микробных сообществ в копролитах разных видов дождевых червей и контрольной почве методом МСТ (мультисубстратного тестирования) выявила их существенные различия. Так, количество потребляемых субстратов и индекс Шеннона в контрольной почве были минимальными, а в копролитах их значения увеличивались. Для копролитов A. rosea оказались характерны высокие показатели количества потребляемых субстратов, разнообразия и выровненности, а также очень высокие – метаболической активности и устойчивости микробной системы. В копролитах L. rubellus, наоборот, эти показатели указывали на дестабилизацию микробной системы. Возможно, негативное влияние связано с выделением этими червями экскретов, снижающих активность микроорганизмов. Содержание двух видов червей вместе нивелировало воздействие каждого вида в отдельности».
Тиунов А. В. «Оценка участия дождевых червей в трансформации органического вещества в дерново-подзолистой почве», 1993:
«Установлено, что дрилосфера собственно-почвенных и почвенно-подстилочных червей, составляющих основу биомассы люмбрицид в дерново-подзолистых почвах, играет столь же существенную роль в формировании биологической активности почвы, как и дрилосфера норных червей».
Биоразнообразие
Б. А. Бызов, Т. Ю. Нечитайло, Б. К. Бумажкин, А. В. Кураков, П. Н. Голышин, Д. Г. Звягинцев «Культивируемые микроорганизмы из пищеварительного тракта дождевых червей», 2007:
«Группировки бактерий, выделяемых из пищеварительных трактов, существенно различаются от группировок из почвы и экскрементов.
Ряд таксонов бактерий оказались общими для различных отделов кишечника A. caliginosa и для кишечников червей различных видов, хотя в целом, состав бактериальных сообществ этих объектов различен. Предполагается, что существуют группировки бактерий, симбиотически ассоциированных с кишечниками.
Сравнение состава бактерий, выделенных из пищеварительных трактов червей Aporrectodea caliginosa, Lumbricus terrestris и Eisenia fetida. В этом эксперименте сравнивали бактериальные группировки всего пищеварительного тракта разных червей. Максимальное количество бактериальных таксонов было выделено из пищеварительного тракта червей A. caliginosa – 43, L. terrestris – 22 таксона, E. fetida – 21».
Цифры ясно дают понять, что биоразнообразие микроорганизмов в кишечнике пашенника в 2 раза выше чем у норника и навозного червя.
Люмбрициды и азот
Терещенко Н. Н. «Эколого-биологические факторы и механизмы ремедиации антропогенно-нарушенных почв» 2007:
«ТНС и мульча одновременно служили приманкой для аборигенных навозных и почвенных червей и способствовали инициации процесса вермикомпостирования непосредственно в корнеобитаемой зоне картофеля».
«Результаты предпринятых сравнительных исследований копролитов дождевых червей, принадлежащих к разным морфо-экологическим группам, позволили сделать заключение об избирательном характере их взаимодействия с микроорганизмами исследованных физиологических и систематических групп.
Основное отличие Красного калифорнийского гибрида от остальных изученных видов навозных (Eisenia fetida киргизской и сибирской популяций) и почвенных червей (Aporrectodea caliginosa) состоит в значительном ускорении темпов аммонификации и минерализации азотсодержащих органических веществ уже на первых этапах переработки ТНС».
«Численность аммонификаторов и микроорганизмов, усваивающих минеральный азот, в копролитах Красного калифорнийского гибрида после первого месяца вермикультивирования значительно (в 6-7 раз) превышала численность данных микроорганизмов в копролитах остальных червей».
«Навозные черви в целом отличаются от почвенных активным стимулированием микробиологических процессов минерализации азотсодержащих органических веществ, а также нитрификации».
«Почвенные черви, напротив, способствуют усилению микробной деструкции лигноцеллюлозного комплекса органического субстрата. Кроме того, почвенные черви являются активными стимуляторами несимбиотической азотфиксации, в десятки раз превосходя по этому показателю навозных червей».
«Для ответа на вопрос о степени сходства и различия между исследованными видами дождевых червей по их влиянию на биохимические и микробиологические свойства производимого вермикомпоста была проведена статистическая обработка всего массива экспериментальных данных при помощи дискриминантного анализа. Сравнение копролитов четырех видов червей по признакам накопления биомассы микроорганизмов, интенсивности выделения СО2 («дыхания»), а также активности таких ферментов как нитрогеназа, целлюлаза, полифенолоксидаза и каталаза показало, что все исследованные виды дождевых червей образуют 4 хорошо обособленные кластера, что, в свою очередь, свидетельствует о наличии значимых отличий в направленности и уровне стимулируемых ими биологических процессов в органическом субстрате При этом наиболее обособленную группу формируют почвенные черви – в природных условиях тяготеющие к принципиально иной среде обитания, чем остальные исследованные представители навозных червей».
«Несмотря на то, что данные дискриминантного анализа в целом относят Калифорнийский гибрид в группу навозных червей, на рисунке он все же заметно обосабливается от навозных червей природных популяций: сибирского и киргизского. Наиболее заметным данное отличие становится при анализе микробиологических процессов цикла азота (содержание в копролитах общего и нитратного азота, накопление аминокислот, численность аммонификаторов, нитрификаторов и денитрификаторов), что подтверждает наличие определённых особенностей в характере стимулируемых гибридом микробиологических процессов и является результатом его направленной селекции с целью адаптации к повышенным концентрациям азота в субстрате".
По этим цитатам можно сделать выводы:
1. Разные морфо-экологические группы дождевых червей стимулируют развитее разных микробных группировок в почве.
2. Навозные черви способствуют связыванию внесённого азота, почвенные – азотфиксации из воздуха.
Отличия вермикомпостов разных червей
Мухитова М. Э. «Сравнительная оценка биотрансформации органических отходов видами семейства Lumbricidae», 2010: «… вермикомпосты почвенных (L. terrestris) и компостных червей (E. fetida) по своим характеристикам достоверно не отличаются. Вермикомпосты калифорнийских червей (E. f. andrei) содержат больше гумуса, чем вермикомпосты червей природных видов».
М. Э.Мухитова, Е. В. Титова «Структурообразующая роль червей семейства Lumbricidae в биогеоценозах».
«Каждый из субстратов подвергался биоконверсии представителями семейства Lumbricidae: обыкновенным дождевым червем Lumbricus terrestris; навозным Eiscnia foetidae и его подвидом E.foetidae andrei, который принято называть калифорнийским красным технологическим гибридом.
… Результаты гранулометрического анализа биогумуса, полученного из кроличьего навоза, переработанного калифорнийскими, навозными и дождевыми червями:
… В продукте, продуцированном навозными червями, – чистый биогумус составлял 88,33%, солома – 1,34%, кора – 10,33%.
В продукте, продуцированном калифорнийскими червями, – 87,83% – чистый биогумус, 1,25% – солома, 10,92% – кора.
В продукте, продуцированном дождевыми червями: чистый биогумус – 84,12%; солома –0,88%, кора – 15%.
… Результаты просеивания биогумуса, полученного из овечьего навоза, переработанного калифорнийскими, навозными и дождевыми червями:
В продукте переработки от навозных червей чистый биогумус составлял 94,74%, кора – 5,26%.
В продукте, продуцированном калифорнийскими червями, – чистый биогумус составлял 92,40%, кора 3,09%, солома 4,51%.
В продукте, продуцированном дождевыми червями, чистый биогумус составляет 92,84%, кора – 6,99%, солома – 0,17%.
Результаты просеивания биогумуса, полученного из свиного навоза, переработанного калифорнийскими, навозными и дождевыми червями:
В биогумусе, продуцированном навозными червями, чистый биогумус составлял 93,88%, солома – 1,52%, кора – 4,6%.
В продукте, продуцированном калифорнийскими червями, чистый биогумус составляет 87,68%, солома – 2,69%, кора – 9,63%.
В продукте, продуцированном дождевыми червями, чистый биогумус составлял 79,84%, солома – 15,84%, кора – 4,32%».
В данном отрывке под словами "чистый биогумус» подразумеваются капролиты. И по этому показателю норники уступают технологическим и местным навозным червям, но совсем незначительно.
Тиунов А. В. «Метабиоз в почвенной системе: влияние дождевых червей на структуру и функционирование почвенной биоты»:
«В копролитах L. terrestris и других видов дождевых червей, собранных в полевых условиях, также преобладали мелкие (<0.01 мм2) фрагменты органического материала (Рис. 4.2). Средний размер органических частиц был достоверно выше в копролитах подстилочных червей Lumbricus rubellus (0.098 мм2), чем в копролитах L. terrestris (0.044 мм2) и двух эндогейных видов, Aporrectodea caliginosa и Octolasion tyrtaeum (0.019 и 0.021 мм2, соответственно)".
По этой цитате: чем сильнее измельчены органические остатки в капролитах почвенных животных, тем интенсивнее идёт их дальнейшая биодеструкция, что хорошо для плодородия почвы. Органические частицы в капролитах пашенника в 2 раза мельче, чем у норников и в 5 раз мельче чем у подстилочных.
Водный баланс почвы в зависимости от капролитов разных дождевых червей
Султан А. Исмаил «Вермикология», Лондон 1997:
«Дождевые черви сильно изменяют пористость почвы. Ещё Wollny в1890 году сообщал, что в почве, заселённой дождевыми червями, возрастает величина соотношения объёмов почва/воздух от 8 до 30%. В связи с этим дождевые черви так же влияют на проникновение воды и её водоудерживающую способность почвы. Данные Stockdill (1966)) относятся к доступной влаге в почве (0-30 см) под участками пастбищ в присутствии или отсутствии люмбрицид в Новой Зеландии. Было поставлено несколько экспериментов для изучения влияния червей на физические свойства почв, особенно относительно подвижности воды. Несмотря на множество трудностей в осуществлении этих опытов, они были необходимы и дали полезную информацию для определения функционально важных морфологических изменений (Diestel, 1992). Кроме дождевых червей-анециков Lumbricus terrestris (Joschko с сотр., 1989), виды-эндогеики Apporrectoidea caliginosa и Apporrectoidea rosea (Joschko с сотр., 1992) так же создают водопроводящие каналы, которые эффективны в насыщенных или затопленных водой почвах».
Прусак А. В., Смагин А. В., Костина Н. В., Умаров М. М., Богданова Т. В. «Гидрофизические свойства и биологическая активность копролитов дождевых червей разных эколого-трофических групп»;
"В лабораторных условиях нами проводился модельный эксперимент по инкубации дерново-подзолистой легкосуглинистой почвы с целью дифференцированной оценки влияния культур дождевых червей разных эколого-трофических групп: почвенно-подстилочных (Lumbricus rubellus) и типично почвенных (Aporrectodea rosea) на гидрофизические свойства и биологическую активность почвы.
… Уже после десятидневного периода инкубации произошли заметные изменения гидрофизических свойств копролитной почвы. Независимо от видового состава червей в сосудах, статистически значимо увеличилась водоудерживающая способность почв в сравнении с контрольными образцами, увеличилась равновесная влажность и влагоемкость, что в целом оказывает положительное влияние на физические свойства почв».
Из цитат становится понятно, что на проникновение воды в почву и водоудерживающую способность почвы одинаково благоприятно влияют как среднеярусные черви, так и черви питающиеся слаборазложившимися органическими остатками – подстилочные и норники.
Дата добавления: 2019-09-08; просмотров: 376; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!