Характеристика медленных колебаний основной активности.
Интенсивность медленных колебаний основной активности не во всех отделах коры одинакова. У кошек она сильнее всего в центральных и теменных долях, слабее —в лобных и затылочных (Беритов и сотр., 13; Цкипуридзе, 14) (рис. 82). Та же закономерность наблюдается у обезьян (Беритов и Гедеванишвили, 15) (рис. 88).
При одновременной регистрации основной электрической активности от двух участков коры оказывается, что медленные потенциалы часто совпадают во времени, а иногда и по амплитуде. Эта синхронность лучше всего проявляется при отведении симметричных участков теменно-центральной области. Синхронность лучше выступает при отведении симметричных участков обоих полушарий, чем несимметричных. У человека эта синхронность
153
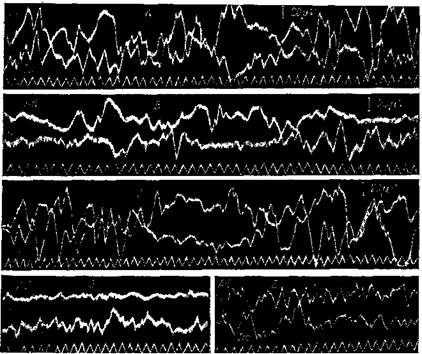
"Рис. 88. Электрическая активность коры большого мозга обезьяны ( Ma c acu s rhesus ) Шлейфный осциллограф. Биотоки регистрируются от двух участков. Биполярное отведение через двухтактные усилители с емкостной связью. А — от симметричных участков центральных долей справа ( C d ) и слева (Cs); В — от симметричных участков затылочных долей справа (Ad) и слева (As); С — от левой центральной доли около височной линии (C's) и около медиальной (Cs); D — от правой затылочной доли (Ad) и от правой лобной доли ( Dd ) в период затишья; Е — те же участки, что и в опыте D , но в период усиленной деятельности (Беритов и Гедеванишвилл)
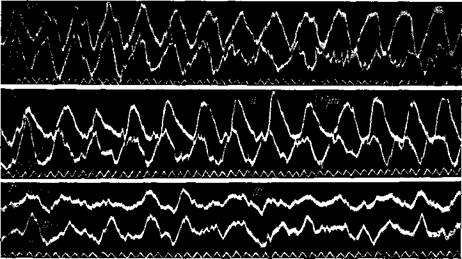
Рис. 89. Биотоки коры большого мозга человека
Регистрация шлейфным осциллографом. Продольное биполярное отведение корковых биотоков от кожной поверхности черепа. Глаза закрыты. I — биотоки от симметричных y4acTKOBj затылочных областей; II — то же ст теменных; III — то же от задней половины лобных долей
выражена лучше, чем у животных. Обычно синхронные колебания однозначны, но в определенных случаях они могут быть антагонистичными (рис. 88, 89).
Медленные колебания основной активности имеют у животных наибольшую амплитуду в теменно-центральных областях коры и ослабевают впереди и позади этого участка; таким образом, эта область является доминирующей в отношении медленных колебаний основной активности (рис. 92).
Характеристика альфа-ритма.
Как уже говорилось, альфа-ритм характерен для человека, но регулярные медленные колебания наблюдаются и в коре больших полушарий животных; это бывает не только при наркозе, но временами и у нормальных животных в бодрствующем состоянии (Цкипуридзе, 14).
У большинства нормальных людей альфа-ритм ясно выражен. Аъ'литуда альфа-волн у нормальных людей не превышает 50—100 мк в, но у сравнительно небольшого числа (10 %) альфа-ритм не возникает. Отсутствие альфаритма часто сопровождается вообще очень низкой амплитудой электрических колебаний без какого-либо доминирующего ритма, при этом у таких людей не обнаруживают каких-либо отклонений от нормы (Адаме, 16). С другой стороны, у некоторых людей альфа-ритм обнаруживает необычайную стойкость, сохраняясь в условиях, при которых обычно он исчезает. При общем наркозе в начале наркотизации (хлороформ, эфир, этилен) альфа-ритм исчезает, уступая место ритму 20—30 в сек.
У каждого человека частота альфа-ритма стойко удерживается с колебаниями + 0,5 цикла в течение многих лет жизни — от полового созревания до старости. Значительные изменения частоты могут быть вызваны ядами нервной системы, когда появляются признаки интоксикации, так прием 100 г алкоголя может привести к изменению частоты альфа-ритма с 10 до 9 в сек.; вещества, вызывающие галюцинации, могут вызвать учащение альфа-ритма с 10 до 12 в сек. (Уолтер, 6). Распределение частот альфа-ритма у большой группы людей следует нормальной Гауссовской кривой, но чаще всего наблюдается частота 10. Анализ альфа-ритма с помощью специальных анализаторов устанавливает, что у каждого индивидуума альфа-волна имеет сложный характер и содержит несколько компонентов из полосы частот 8—13 в сек. При полном покое в отсутствие умеренной работы происходит усиление медленных компонентов, например 9 в сек.; при усиленной умственной рабо те усиливаются более быстрые компоненты, например 11 в сек. (Уолтер, 6).
Установлено, что альфа-ритм не имеет отношения к психике, интеллектуальному развитию и вообще к каким-либо специфическим функциям мозга человека (Лемер, 17; Беритов, Бакурадзе и Дзидзишвили, 18; Генри и Кнотт, 19; Карр, Поллак, Финк, 20). Это важное заключение в равной степени касается и бета-ритма.
Характер альфа-ритма не связан и с полом. Но, очевидно, характер альфа-ритма связан с типом высшей нервной деятельности человека, со степенью возбудимости ЦНС. Люди с медленным альфа-ритмом выполняют психо-технические тесты медленно, люди с быстрым альфа-ритмом — быстрее (Уолтер, 6). Это согласуется с классификацией, предложенной Беритовым (21), по которой частота 11, 12 в сек. характеризует людей с высокой корковой возбудимостью, частота 9—10—с низкой. Мы увидим ниже, что это имеет, вероятно, основания в свойствах корковых нейронов.
Альфа-ритм лучше всего выявляется в темноте, при закрытых глазах, когда испытуемый лежит спокойно, без напряжения мышц головы и без усиленной умственной работы. При других условиях альфа-ритм проявляется хуже. Открытие глаз, освещение, умственная работа и волнение мешают проявлению альфа-ритма. Если при открытых глазах на свету альфаритм отсутствует, то закрывание глаз приводит обычно к его появлению со скрытым периодом около 0,3—0,5 сек.; в первые секунды частота его на 0,5—1,5 цикла выше, чем потом (Адамович, 22). Если альфа-ритм присутствовал в ЭЭГ до закрывания глаз, то после закрывания он зна-
155
чительно усиливается (рис. 87). Так же действует темнота: если потушить свет при открытых глазах, альфа-волны усиливаются, если их не было до затемнения, то они появляются; скрытый период появления альфа-ритма в этом случае длиннее — более 1—2 сек. (рис. 87). Альфа-волны периодически то усиливаются, то ослабевают через каждые 1—2 сек., но у некоторых лиц они возникают группами, с непостоянным интервалом времени между ними.
Одним из характерных явлений, обнаруживаемых при регистрации ЭЭГ строго симметричных участков обоих полушарий, является синхронность электрических колебаний (Эдриан и Ямагива, 23; Беритов, 3); из 1056 пар альфа-волн затылочной области 50% синхронны в пределах 1 мсек., а в пределах 10 мсек.— 75% (Гарут и Айрд, 24). Временами синхронность альфаволн нарушается, но вновь восстанавливается, т. е. имеется тенденция альфа-волн двух симметричных пунктов коры, утративших синхронность, возвращаться к ней (рис. 89); это указывает на наличие какого-то саморегулирующегося механизма (Гарут и Айрд, 24). В одном полушарии альфа-волны возникают в разных участках коры в разное время: в передних частях альфаволны достигают вершины раньше, чем в затылочной. Характерным для альфа-ритма является сдвиг фаз на 90° при поперечном и продольном отведениях, например, парието-темпоральные электроды могут регистрировать вершину альфа-волн, а парието-окципитальные — нулевой потенциал (Уолтер, 6). При некоторых заболеваниях мозга, в частности при шизофрении (Дзидзишвили, 25), синхронность альфа-волн может резко нарушаться.
Регулярный альфа-ритм 8—13 в сек. бывает у человека, но, как уже говорилось, аналогичные медленные колебания временами становятся регулярными и у животных (Цкипуридзе, 14; Кратин, 26), хотя в этих периодических вспышках ритм быстро меняется в больших пределах. Под влиянием барбитуратов медленные потенциалы в коре у животных могут принимать правильный ритм около 10 в сек., но этот ритм нельзя аналогизировать с естественным альфа-ритмом.
Альфа-волны, достигая большой амплитуды, могут становиться двуфазными; при небольшой амплитуде они однофазны. При малой амплитуде альфа-волны сливаются, т. е. не успевает данная волна достигнуть абсциссы, как начинается следующая волна. Слившиеся альфа-волны получили название «эта-волн» (Беритов, Бакурадзе и Дзидзишвили, 18).
Когда альфа-волны двуфазны, они часто имеют характер синусоиды, т. е. состоят из двух противоположных фаз, как бы полуволн одинаковой амплитуды; но при специальном анализе обнаруживается, что в действительности форма альфа-волны совершенно не соответствует синусоиде — переход первой фазы альфа-волны ко второй фазе часто происходит с большей быстротой, чем переход от второй фазы одной альфа-волны к первой фазе последующей; от второй фазы одной альфа-волны до первой фазы другой иногда включается небольшая пауза, проявляется это без какой-либо периодичности. Итак, альфа-волны протекают не синусоидально, вторая фаза альфа-волны имеет самостоятельное происхождение. Есть основание полагать, что положительная фаза связана с преимущественным активированием глубоких слоев коры, отрицательная фаза — с преимущественной активацией поверхностных слоев (Беритов, 21).
Центральным местом возникновения альфа-волн является теменная область коры на границе с затылочной — здесь они обладают наибольшей амплитудой и наибольшей регулярностью. Кпереди и кзади от этой области альфа-волны постепенно ослабевают (Эдриан и Ямагива, 23; Беритов, 3). Признавалось существование также второго центра альфа-волн в лобных долях (Рубин, 27; Лумис и сотр., 28), но это не оправдалось исследованиями других авторов (Беритов и сотр., 3, 13, 18) и, в частности, при отведении непосредственно от обнаженной коры человека (Скарф и Рам, 29). У кошек участок, где возникает альфа-подобный ритм наибольшей интенсивности и наиболее регулярного характера, расположен в сенсомоторной области, около крестовидной борозды (Кларк, и Уорд, 30; Цкипуридзе, 14).
156
У ряда лиц электрическая активность центральной (роландической) области в виде ритма 8—13 в сек. бывает выражена не хуже, а иногда и лучше, чем альфа-ритм теменно-затылочной области (Адамович, 22). В то время как теменно-затылочный альфа-ритм резко угнетается при зрительных раздражениях, роландический ритм резко угнетается при мышечной деятельности (Гасто, 31). Характерны антагонистические отношения электрической ритмики теменно-затылочной и роландической областей: при полном угнетении затылочного альфа-ритма вследствие зрительного раздражения роландический ритм может сохраниться и даже усилиться. Это явление демонстративно выступает при чтении книги, наоборот, при мышечной работе полное угнетение роландической ритмики может сочетаться с наличием выраженного затылочного альфа-ритма (Ройтбак, Дедабришвили и Гоциридзе, 7). Электрический ритм роландической области угнетается уже при одном только представлении мышечной работы (Гасто, 31), а в затылочной альфа-ритм — при зрительных представлениях (Слаттер, 32). Но при сильных световых раздражениях и в начале интенсивной мышечной деятельности угнетение ритма 8—13 в сек. наступает во всей коре.
Дата добавления: 2019-07-17; просмотров: 355; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!