Центробежная связь зрительной коры с сетчаткой.
Как уже указывалось, зрительная воспринимающая область связана с сетчаткой также центробежными путями. Опыты с разрушением участков зрительной коры показали, что дегенерированные нисходящие волокна можно проследить до самой сетчатки. Часть этих волокон оканчивается в наружном коленчатом теле, в переднем двухолмии и в других ядрах мозга, а часть достигает сетчатки (Школьник-Яррос, 37; Крэг, 38) (рис. 50). Р. Кахал (5) подробно исследовал центробежные волокна, приходящие по зрительному нерву в сетчатку. Они проходят через волокнистый слой сетчатки, достигают слоя амакриновых клеток. По Поляку (39), эти волокна ( t — на рис. 51) заканчиваются на особого рода биполярных клетках (г), относимых к ассоциационным элементам сетчатки. Один отросток этих клеток образует синапсы на основании колбочек ( b ), a другой контактирует с отростками других биполяров ( h f ) и ганглиозными клетками зрительного нерва.
При активации зрительной коры, когда происходит восприятие или репродукция образа внешнего мира, должны также активироваться центробежные пути с включением означенных биполярных клеток сетчатки. Характерно, что при электрическом раздражении зрительной области в одном
78

Рис. 51. Упрощенная схема структуры сетчатки, демонстрирующая распространение импульсов от фоторецепторов к другим частям сетчатки, к головному мозгу и назад от мозга к сетчатке (направление покавано стрелками)
а, b — фоторецепторы — палочки и колбочки, где зарождаются нервные импульсы при воздействии Света (раздражается только левая группа фоторецепторов); с—горизонтальная клетка, которая проводит импульсы к окружающим колбочкам и палочкам; е-/, h , d — разнообразные биполярные клетки, которые проводят импульсы к ганглиозным клеткам (m, n , s , p , о); i —биполярная клетка, разновидность амакрииной клетки, которая получает импульсы как от фоторецепторов и проводит их центрипетально к ганглиовным клеткам, так и из мозга по центрифугальным волокнам (() и проводит назад к фоторецепторам (Ь); I —амакриновая клетка, которая, возможно, перехватывает часть биполярных импульсов и передает их на соседние участки: m, n, s , р, а — ганглиозные клетки, которые получают импульсы от центрипетальных биполярных клеток и передают их к головному мозгу через зрительный нерв (из Поляка)
полушарии человек слепнет на противоположный глаз (Пенфилд и Джаспер, 29). Возможно, что при некоторых силах раздражения в коре возбуждается столько центробежных нейронов, и сообразно в сетчатке активируется такая масса амакриновых и биполярных клеток, что в силу рефрактерной фазы они перестают реагировать на адекватные световые раздражения. Нисходящие влияния от коры на сетчатку показаны и другими исследователями (40, 41).
Надо полагать, что активирование центробежной системы играет важную роль в восприятии и создании образов внешнего мира. А так как центробежная система стимулирует в сетчатке ту же систему нейронов, как и адекватное раздражение глаза, то можно предположить, что основное функциональное назначение центробежных путей к сетчатке заключается главным образом в локализации или проецировании зрительных образов внешних объектов в тех же участках внешней среды, где эти объекты находились в момент их восприятий.
79
Воспринимающая область слуха.
Вопросы сравнительной физиологии слухового анализатора выходят за рамки данного раздела. С ними можно познакомиться в полном новейшем обзоре Шварцкопфа (42). Хотя внутреннее ухо птиц и обладает базилярной мембраной, оно значительно отличается по анатомии от внутреннего уха млекопитающих (43). Интересно отметить, как показали Бурд и Расмуссен (44), что лагенарные волокна (lageпае — терминальная часть улитки позвоночных, кроме млекопитающих) идут в вестибулярную систему, в ретикулярную формацию и к ядрам мозжечка. Позднее мы покажем, какую роль выполняют указанные связи у млекопитающих. Электрофизиологически у птиц исследованы акустические проекции и вызванные потенциалы продолговатого мозга, базальных ядер и фронтального неостриатума (Ильичев, 45).
До последнего времени основным методом исследования анатомических границ слуховой функции, как и других воспринимающих функций, являлось удаление определенных частей коры мозга. Этим путем было установлено, что слуховая функция у высших позвоночных животных осуществляется главным образом в области экстосильвиевой извилины в височных долях, (рис. 46, В). Но данный метод не является точным, ибо собаки и кошки лишенные всей коры, реагируют на звуки. Даже децеребрированная кошка проявляет рефлексы на звуковые раздражения (Форбс и Шеррингтон, 46). У кошек после |удаления височной доли в обоих полушариях сохраняется способность локализации направления звуков. Спустя 6—7 недель после полного удаления новой коры кошка идет по направлению к звуку (Орджоникидзе и Нуцубидзе, 47).
В настоящее время пределы воспринимающих областей в коре больших полушарий устанавливаются электрографическим путем. Раздражается рецептор и исследуется изменение электрической активности в том или другом участке коры мозга. Этим путем было показано, что у кошки и собаки воспринимающая функция слуха осуществляется в области верхней части сильвиевой и средней части эктосильвиевой извилины, а также передней и задней части эктосильвиевой извилины (Корнмюллер, 48; Бремер и Доу, 49; Тунтури, 50). Детальная топография воспринимающих слуховых областей была получена при применении звуковых раздражений: тона, щелчка, а также при локальном электрическом раздражении кохлеарного нерва в базальном среднем и апикальном завитках улитки (Булей, 51).
Восприятие звуков разной высоты производится разными участками коры и эти участки располагаются сообразно с высотой звуков (Ларионов, 52). Путем изучения корковых потенциалов при разных звуках было установлено, что в средней части эктосильвиевой извилины звуки более высокой частоты воспринимаются передними ее участками, а низкие — задними; причем последовательные октавы располагаются друг за другом вдоль коры через более или менее одинаковые интервалы (Тунтури, 50).
Но при образовании условных рефлексов на звуки разной высоты в условиях двустороннего повреждения средней части эктосильвиевой извилины эта дифференциация полностью не исчезает. Она только становится более грубой (Мосидзе, 53).
В высшей степени вероятно, что средняя часть эктосильвиевой извилины имеет отношение главным образом к восприятию звуковых раздражений и является первичной зоной. Что же касается анализа и синтеза их, в особенности сохранения воспринятых звуков в памяти, то эти функции должны осуществляться главным образом сильвиевой и супрасильвиевой извилинами, которые составляют вторичную зону.
У человека слуховая область находится в середине верхней височной извилины и в так называемой геншеновской извилине, которая начинается в глубине сильвиевой борозды (Геншен, 26).
При поражении корковой слуховой зоны у человека (поля 41, 42, а также 22) нарушается восприятие низких звуков при поражении более передних отделов, а восприятие высоких — при поражении каудально расположенных
80
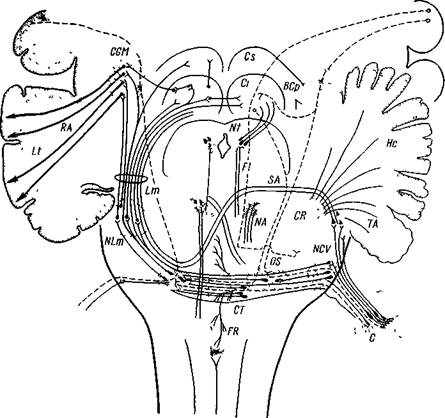
Рис. 52. Схема восходящих чувствительных п > тей слухового нерва (слуховой анализатор)
Дана схема В Бехтерева, видоизмененная А Микепадзс на основании современных литературных данных На ней даны как кортикопетальные пути, так и кортикофугальные (штрихами) L t — Lobus temporalis, ЛА — Radiatio acustica, C O M — Corpus gemculatum mediale, Lm — Lemniscus mediahs, N Lm — Nucleus lemniscus mediahs, Cs — Colliculus superior, Сг — Colhculus inferior, Nt — Nucl n trochlearis, F l — Fasciculus longitudmahs mediahs 8 A — Stria acustica, NA — Nucl. n abducens, OS — Oliva superior, CT — Coipus trapezoideum, FR — Formatio reticularis, B Cp — Brachu conj post , СД — Corpus restiformis, 1\ LV —Nucl cochlcaris ventralis, He—Hemisphern cerebelli! С — Cochlea, ТА — Tuber acusticum
отделов. Во вторичной корковой слуховой зоне, наоборот, при поражении более передних отделов страдают высокие звуки, а каудальных — низкие (Литвак, 54).
Афферентные пути от слухового рецептора до коры больших полушарий проходят главным образом через специфические ядра (рис. 52) (55). Улитковое ядро продолговатого мозга (nucleus cochlearis) — кохлеарный комплекс может быть разделен на три части (Авксентьева и др., 56): ретику лярную (заднее вентральное ядро), ядерную (переднее вентральное ядро — основное у человека, Циммерман, 57) и корковую (слуховой бугорок). Его связи с ретикулярными элементами обеспечивают выход сигналов на моторную систему. В комплексе верхних олив имеется самый низкий уровень конвергенции сигналов с левого уха и правого уха. Существенную роль здесь выполняют нейроны акцессорного ядра (Мушегян, 58).
Афферентные пути от слухового рецептора частично проходят и через мозжечок. Здесь они прерываются в области lobus simplex и tuber vermis (Снайдер и Стоуел, 59). Сложный путь через мозжечок играет важную роль в проецировании и удержании на долгое время воспринятых звуков. Это видно из того, чю после удаления мозжечка нарушается способность к удерживанию в памяти направления воспринятых звуков. Нормальная собака с зак-
81
рытыми глазами может прямо пойти к месту стука миски, из которой она получает еду. Это она сделает не только во время стука, но и через несколько минут после стука, т. е. животное воспринимает не просто звук, но и его направление и место источника звука. После удаления мозжечка оно продолжает воспринимать стук миски, ибо поворачивает голову в сторону звука и даже пойдет к месту стука, если он все время продолжается. Но по прекращении стука животное не в состоянии пойти к источнику звука — к пище; оно начинает поисковые движения в том самом месте, где застает его прекращение стука. Видимо мозжечок через специфическую систему ствола мозга или через неспецифическую систему — ретикулярное образование влияет облегчающим образом на слуховую воспринимающую область. От этого она становится способной не только воспринимать звуки, но и удерживать образы их на долгое время (Беритов, 60).
Афферентные пути отдают также коллатерали к ретикулярному образованию среднего мозга (см. рис. 52) и через этот неспецифический путь слуховые импульсы достигают слуховой коры (Галамбос, 61), а также других областей коры, особенно вестибулярной (Юнг и др., 36) (см. рис. 49—II).
Большой фактический материал получен в последнее время с помощью микроэлектродной регистрации активности отдельных элементов в различных звеньях слухового анализатора и в слуховой коре. При исследовании влияния параметров стимуляции было показано, что один и тот же стимул может вызвать разряд в различных элементах (Герштейн и Киянг, 62). Как нами было уже отмечено, одиночные нейроны в различных звеньях анализатора отличаются разнос оразием реакций. В слуховой коре имеются нейроны по «on — off» типу (Кацуки, 63), или отвечающие только на смену частоты звука (Эванс и Вайтфилд, 64).
Полагают, что вертикальная организация слуховой коры отличается от таковой соматосенсорной коры (Маунткэстл, 97) и зрительной (Хьюбель и Визел, 34, 35).
От каждого уха звуки воспринимаются в обоих полушариях, но более сильно — в противоположном полушарии. Наличие конвергенции сигналов от левого уха и правого уха объясняется анатомическими данными — наличием нескольких уровней перекреста. Метод одновременного или раздельного предъявления на оба уха равных по интенсивности звуковых сигналов (щелчков) позволяет по соотношению электрических реакций отдельных нейронов при ипси-, контралатеральном и при бинауральном раздражении показать конвергенцию во всех звеньях и, естественно, в слуховой коре большого мозга (Альтман, 65). Однако в настоящее время нет полной и достаточно экспериментально обоснованной теории бинаурального слуха.
Слуховая воспринимающая область также имеет центробежную связь с рецептором — кортиевым органом (рис. 52). Эфферентные пути прослежены морфологически и подтверждены в физиологических экспериментах. После разрушения коры в слуховой зоне и в окружавших ее областях дегенеративные кортикофугальные волокна прослеживаются в медиальном коленчатом теле,в промежуточном, среднем мозге и мосте (Десмет и Мехелс, 66; Эрнандец-Пеонидр., 67). Кроме того, показано, что контралатеральное удаление слуховой зоны снимает ранее наблюдавшийся эффект уменьшения потенциала возбуждения слухового нерва у кошки (Альтман, 68).
При повреждении же внутри продолговатого мозга в области ножек оливы дегенерированные эфферентные волокна были прослежены до кохлеарного ядра и далее до кортиева органа (Расмуссен, 69). Где эти последние контактируют, еще не известно. Они настолько тонки (микрон и еще меньше), что пока не удается проследить их до конечных разветвлений. Однако известно, что при возбуждении этих волокон слуховые волосковые клетки перестают реагировать на адекватные слуховые раздражения (Галамбос, 70). Очевидно, центробежные волокна возбуждают те самые чувствительные элементы, на которые действуют звуковые волны. В последнее время показано, чта при раздражении эфферентных путей Расмуссена наиболее эффективно
82
подавляются ответы на низкие по частоте и слабые по интенсивности звуковые раздражения (Зомер, 71); при выключении их прокаином (Лейбрандт, 72) наблюдали нарушение адаптации на ритмически подаваемый тон.
Дата добавления: 2019-07-17; просмотров: 369; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!