ГЛАВА IV. ВОСПРИНИМАЮЩАЯ ФУНКЦИЯ КОРЫ БОЛЬШОГО МОЗГА
Воспринимающие области в коре большого мозга.
Возбуждение каждого рецептора неизбежно оказывает свое влияние и на кору больших полушарий. Для каждого рецептивного органа в коре млекопитающих существует определенная воспринимающая территория (Феррье, 1; Мунк, 2). Причем у низших млекопитающих, например у кролика или кошки, каждая функция занимает значительную часть мозговой поверхности, перекрывая одна другую, но в каждой воспринимающей территории имеется одна оптимальная область. После удаления этой области данная функция страдает очень сильно, но полностью не исчезает.
У высших млекопитающих каждая из воспринимающих функций все более и более сосредоточивается в соответственной оптимальной области, вследствие чего значительная часть мозговой коры освобождается от воспринимающих функций. Так, у высших обезьян почти l / s всей коры, а у человека 2Д всей коры не играют существенной роли в отношении воспринимающей функции. Однако по клиническим наблюдениям, даже у человека в некоторой, очень малой степени, воспринимающая функция сохраняется во всех отделах коры мозга (Монаков, 3; Гольджтейн, 4).
В каждой воспринимающей области выделяют ядерную, или первичную зону, где находится главная масса сенсорных звездчатых нейронов. Эти нейроны сосредоточены главным образом в четвертом слое, где оканчиваются соответствующие афферентные пути. У высших млекопитающих, как у обезьяны и человека, четвертый слой очень широк и делится на несколько подслоев. Мы уже говорили, что восприятие внешнего мира, создание образов или конкретных представлений происходит благодаря активации сенсорных явездчатых нейронов афферентными импульсами. Вокруг каждой первичной зоны находятся так называемые вторичные зоны. Здесь сенсорных звездчатых клеток значительно меньше, поэтому четвертый слой очень узкий. Взамен сильно развит третий слой. В этом слое, а также в четвертом сосредоточены главным образом вставочные и ассоциационные пирамидные нейроны. Как будет показано ниже, значение вторичных зон заключается в основном в том, что ассоциационные пирамидные нейроны этих зон участвуют в сохранении образов внешнего мира, возникающих в первичной зоне.
Каждая воспринимающая область связана с рецептором не только путем центростремительных афферентных путей, исходящих от рецептора и прерывающихся один или несколько раз в стволе мозга. Она связана с рецептором также центробежными нервными путями и тоже с перерывами в стволе мозга. На эти центробежные пути обратили внимание еще Кахал (5) и Бехтерев (6). В последнее время они хорошо изучены в отношении мышечных рецепторов, и центробежные нервные волокна, оканчивающиеся на мышечных веретенах, были обозначены как гамма-система (Беритов, 7; Меттьюс, 8; Гурфинкель и др., 9). Сигналы, приходящие по гамма-эфферентам, вызывают
70
укорочение полярных частей интрафузальных волокон, вследствие чего либо появляется афферентный разряд, либо изменяется (учащается) разряд, вызванный предшествующим растяжением мышцы.
На приводимых ниже рисунках зрительного и слухового анализаторов центробежные пути изображены штриховыми линиями. Предполагается, что кора мозга этими центробежными путями регулирует деятельность самого рецептора, облегчая или угнетая его реагирование на внешнее адекватное воздействие (Гранит, 10; Галамбос, 11; Хагбарт, 12). Но наряду с общей функцией центробежная система нейронов, вероятно, выполняет еще специфическую функцию проецирования образов воспринятых объектов, подобно тому, как это бывает при самом восприятии. Впервые еще В. М. Бехтерев (9) предположил эту функцию центробежной системы.
Подробнее мы остановимся на этом при обсуждении отдельных воспринимающих областей.
Воспринимающая об л асть зрения. Вопрос о специфическом представительстве у примитивных позвоночных при исследовании вызванных потенциалов, по-видимому, решен отрицательно, т. е. никаких изменений потенциалов на поверхности переднего мозга у них не обнаружено (Карамян и др., 13). Однако у амфибий и рептилий такие теленцефалические ответы зарегистрированы. Уточнения, полученные в последнее время (Карамян, 14; Наута, 15), показали, что локализация зрительных и соматических ответов занимает всю площадь примитивной «общей коры», причем, например у варанов, в «коре» превалируют соматические, а у черепах зрительные проекции (14). В последнее время в лаборатории Г. Д. Смирнова детально исследованы особенности нейронной организации зрител ь ной покрышки у низших позвоночных, причем по сравнению с высшими позвоночными обнаружен малый процент нейронов с фоновой активностью. Большинство из них отвечает только при световом раздражении (Смирнов с сотр., 16—19).
У птиц воспринимающая функция представлена не только в коре больших полушарий, но и в подкорковых центрах, промежуточном мозге (Шрадер,20; Беритов,60). У птиц, в частности у голубей, большой мозг безусловно владеет зрительной функцией, ибо с удалением коры происходят некоторые расстройства в области зрения: поведение птицы свидетельствует об отсутствии узнавания знакомой среды. Но голубь с удаленной корой избегает препятствий и при полетах хорошо различает предметы, на которые он садится. Анатомические исследования показали, что у голубя имеется текто-таламо-стриарный путь: в наиболее глубоком клеточном слое tectum opticum начинается хорошо очерченный путь к п. rotundus thalami и далее путь к ectostriatum (Картен и Ревзин, 21). Однако зрительная система в ectostriatum и слуховая в striatum у голубя не имеют гомологов в переднем мозге млекопитающих.
На зоологической лестнице позвоночных корковая зрительная область впервые отмечается у птиц. Она занимает поверхность полушарий впереди и сверху мозговых ножек и прикрывает мозговой желудочек.
Кора большого мозга млекопитающих (кошки, собаки) обладает зрительной функцией в полной мере. На основании опытов школы И. П. Павлова можно утверждать, что для высшего анализа и синтеза зрительных раздражений служит определенная область в затылочной части коры, а именно в ее верхней, задней и медиальной поверхностях (рис. 46).
По исследованиям Мунка (2) на собаках и других высших позвоночных, а также на человеке, зрительная область одного полушария связана с сетчаткой обоих глаз. Внутренняя, носовая часть сетчатки связывается с корой противоположного полушария. Наружная, височная часть сетчатки связывается с полушарием своей же стороны (рис. 47). От центральной, макулярной части сетчатки волокна идут в соответствующее и противоположное полушарие (Ринг, 22). Перекрещивание зрительных путей происходит в хиазме зрительных нервов. У низших млекопитающих, а также у птиц, рыб я амфибий имеется полное перекрещивание зрительных волокон. Поэтому
71
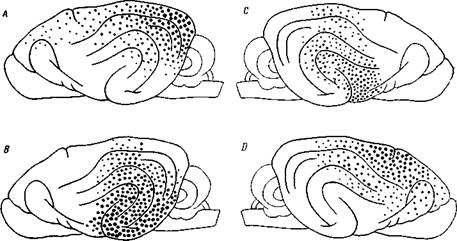
Рис. 46. Топография воспринимающих областей у собаки
А — зрительной, В — слуховой, С — вкусовой, D — кошно-мышечной. Точки обозначают соответствующие поверхности. Жирными точками обозначены участки мозга, обладающие наибольшей воспринимающей функцией — первичная зона (Лучиани)
у этих животных сетчатка одного глаза связывается только с противоположным полушарием.
Если у новорожденных высших позвоночных животных удалить глазные яблоки, то зрительный нерв и его продолжение в мозге перерождаются; перерождаются также наружное коленчатое тело, подушка и передние холмы четверохолмия; в зрительной области затылочных долей замечается остановка развития. В первичной зоне зрительной области перерождаются главным образом звездчатые клетки четвертого слоя (Дзидзишвили, 23).
Экстирпация зрительной области у молодых животных приводит к перерождению наружного коленчатого тела, подушки и передних четверохолмий; несколько позднее дегенерирует зрительный нерв (Монаков, 3). У обезьян на 4—5-й день жизни при экстирпации только 17 поля (и небольшой части 18 и 19) в коленчатом теле сохраняется лишь очень тонкий слой клеток во внутренней части переднего отдела; подушка и передние бугры в этом случае не содержат изменений (Джалагония и др., 24).
У обезьян, а также у человека, зрительная область локализуется в верхнем отделе затылочной доли (рис. 48). Одностороннее разрушение ее вызывает частичную слепоту на оба глаза. Согласно исследованиям Минковского (25), у животных существует более или менее выраженное территориальное соответствие между сетчаткой и корой мозга. Так, он находит, что передняя часть зрительной воспринимающей области связывается главным образом с нижней частью сетчатки, но в то же время каждый ее участок имеет множество связей со всей воспринимающей областью. К аналогичному заключению пришли и относительно представительства сетчатки в коре человеческого мозга (Гешпен, 26).
Наличие зрительной воспринимающей области в затылочной доле коры было доказано также опытами и наблюдениями при хирургических операциях на людях. Кору больших полушарий обнажали в целях удаления заболевших участков мозга, а для установления очага заболеваний применяли электрическое раздражение. Оно производилось в отсутствие наркоза и больные активно участвовали в установлении характера ответных реакций на раздражение. При этих условиях раздражение зрительной области вызывало ощущение света, темноты и различных цветов. При раздражении же других областей коры ничего подобного не ощущалось (Пенфилд и Джаспер, 27).
72
В зрительной области первичная зона у высших млекопитающих, как уже говорилось, находится в 17 поле по Бродману. Здесь слой IV развит сильнее других слоев и в нем звездчатых клеток с околоклеточной аксонной сетью неизмеримо больше, чем в слое IV вторичной зоны, которая находится в 18 и 19 полях. В этих полях сильно увеличен третий слой и в нем имеется большее количество вставочных и ассоциационных нейронов, чем в других слоях. На рис. 61 даны соотношения нейронов в первичных и вторичных зонах зрительной коры, связь их с афферентными таламическими волокнами и затем их двусторонняя связь коры с сетчаткой (Школьник-Яррос, 28). Далее известно, что с филогенетическим развитием площадь вторичных зон 18 и 19 полей сравнительно увеличивается больше, чем первичной зоны (Филимонов, 29). Поля 18 и 19 филогентически более молодые структуры, чем поле 17 (Поляков, 30). Онтогенетически поле 17 созревает раньше, чем поля 18 и 19 (Преображенская, 31). Было сделано заключение, что
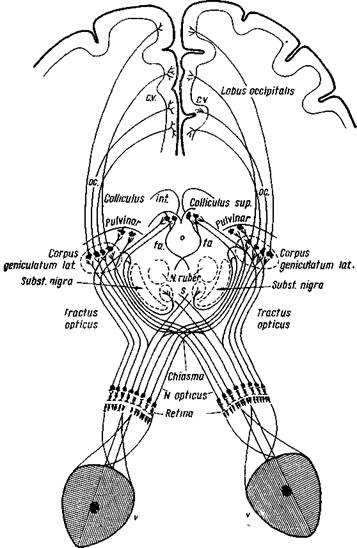
Рис. 47. Зрительный анализатор
V — оптическая система глаза; С. У. — зрительная область коры больших полушарий; О.С. — подкорковые зрительные пути; fa — волокна, переходящие с четверохолмия в fasciculus praedorsalis; s —волокна из хиазмы, идущие ко дну третьего желудочка (Бехтерев)
73
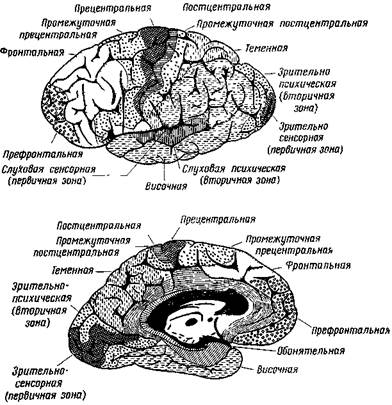
Рис. 48. Воспринимающие области в коре больших полушарий человека
Показаны первичные и вторичные зоны зрительной и слуховой областей и затем все остальные корковые области
в то время как 17 поле необходимо для зрительного восприятия внешнего мира, 18 и 19 поля служат для высшей интеграции зрительной корковой деятельности, связанной с ее психической деятельностью, прежде всего в виде сохранения и репродукции образов внешнего мира (Беритов, 32).
Функциональные особенности первичных и вторичных зон в зрительной области хорошо были выявлены также путем физиологического исследования. Так, например, Бонин, Гэрол и Мак Куллоч (33) производили у обезьян локальное отравление стрихнином одного из полей и наблюдали при этом изменения электрической активности в других полях. Оказалось, что при отравлении поля 17 судорожные разряды появлялись в этом поле и еще в соседнем, 18 поле; при отравлении 18 поля они возникали в 17 поле того и другого полушария и еще в 19 поле соответствующей стороны, а при отравлении 19 поля — только в этом поле. Отсюда авторы заключили, что 18 поле получает импульсы только из 17 поля, а со своей стороны посылает импульсы как обратно к 17 полю, так и к 19 полю той же стороны я еще к 18 полю противоположного полушария; из 19 поля стрихнинные разряды не переходят на клетки других полей; видимо, ассоциационные нейроны из 19 поля оканчиваются главным образом на дендритах других полей, и поэтому авторы наблюдали в этих полях угнетение электрической активности.
Из этих физиологических наблюдений можно сделать одно очень важное заключение: 17 и 18 поля тесно связаны между собой обоюдосторонне я поэтому при раздражении сетчатки они активируются как единое образование, производящее зрительные образы внешних объектов. Поле 19, активируемое в это время из 17 поля, должно быть, производит общее торможение
74
|
|
коры и тем охраняет локальность и целостность данного зрительного образа от одновременного воздействия других объектов.
В поле 19 имеются звездчатые клетки, к которым также подходят афферентные волокна из сетчатки через подушку (см. рис. 61). Каково же значение этого нервного субстрата Из экспериментов известно, что после удаления первичной зоны зрительного анализатора условные рефлексы на световые раздражения не исчезают. Животное теряет предметное зрение, не дифференцирует световые фигуры, но образует рефлекс на свет и дифференцирует его от темноты. Вот эта способность ощущать вообще свет в данных условиях, может быть, зависит от тех зрительных волокон, которые прерываются не в коленчатом теле, а в подушке, и активируют афферентные нейроны, заканчивающиеся в 19 поле на звездчатых клетках IV слоя.
Указанная роль волокон, проходящих через подушку, подтверждена и опытами Хананашвили (см. в гл. V, 24, 25) с разрушением наружных коленчатых тел и последующим выпадением предметного зрения при сохранении восприятия освещенности.
Осциллографическими исследованиями электрической активности коры мозга было также установлено, что раздражение сетчатки активирует первым долгом 17 поле. Здесь возникают ответные первичные потенциалы.
По исследованию Юнга (36), в 17 поле находятся пять типов корковых клеток, реагирующих на зрительное раздражение. Все они обнаруживают спонтанную активность в виде периодического наступления пиковых разрядов. Нейроны типа А вообще не отвечают на освещение, на вспышку света, ни учащением, ни торможением спонтанного ритма. Нейроны типа В реагируют на каждое освещение сетчатки: в период освещения разряды учащаются, а при затемнении наступают реже, т. е. тормозятся. Тип С — разряды тормозятся как от света, так и от темноты. Тип D — нейроны реагируют только при затемнении. Наконец, нейроны типа Е реагируют как на освещение, так и на затемнение. Только при затемнении они активируются с коротким скрытым периодом (рис. 49—/).
По этим опытам видно, что когда определенные нейроны типа В, отчасти типа Е, возбуждаются от света, другие нейроны в это время более или менее тормозятся. Точно так же, когда по выключении света активируются нейроны типа D , а также некоторые нейроны типа Е, в это время нейроны типа В тормозятся.
Юнг не дает объяснения происхождению этих реципрокных отношений в активации корковых нейронов. Он лишь мельком ссылается на концепцию Беритова о координирующей роли дендритных потенциалов и промежуточных нейронов коры, как пригодную для обсуждения описанных явлений. Мы думаем, что на самом деле, согласно нашей концепции насчет происхождения торможения, можно сделать первый шаг к объяснению этих явлений взаимоотношения корковых нейронов.
При внеклеточном микроэлектродном способе отведения обычно отводятся сравнительно большие пирамидные клетки. В опытах Юнга, вероятно, отводились в зрительной коре не сенсорные звездчатые клетки, а пирамидные клетки тех нейронных кругов, которые активировались при освещении и затемнении находящейся перед глазами кошки обстановки опыта. Значит, нейроны типа А, В, С, D и Е, должно быть, являлись пирамидными. Хотя эти нейроны находятся в коре в близком соседстве, они могли входить в разные нейронные круги. Поэтому при преимущественном возбуждении одного круга, например круга освещения, с активацией соответствующих сенсорных звездчатых нейронов, должны были активироваться дендриты пирамидных клеток другого круга, т. е. затемнения, и вызывать в них анэлектротоническую блокаду клеточных синапсов и тем производить торможение соответствующих пирамидных клеток, а значит и всего данного нервного круга. Наступающее по прекращении освещения усиленное активирование клеток круга затемнения должно было происходить от повышения возбудимости в их синапсах вследствие прекращения анэлектротона.
75

Рис. 49—1 . Схема электрической реакции нейронов зрительной коры на освещение и затемнение и их отношение к возбуждению с'рецептора и электроэнцефалограмме
Верхняя кривая показывает потенциал сетчатки при отведении микроэлектродами от слоя колбочек и палочек, а нижняя кривая — от поверхности коры 17 поля при включении и выключении света. Л — Е — схематические разряяы А — Е типов нейронов: А — нейрон совсем не реагирует; В — активизируется при свете и тормозится при выключении света; С — тормозится как при освещении, так и при затемнении; D — тормозится при свете и активируется при затемнении; Е — сначала торможение, затем активация, а после затемнения активация с самого начала (Юнг)
Новейшие исследования Хыобела и Визела (34, 35) по определению рецептивных зон в зрительной коре показали, что в 17 поле преимущественно простые зоны, причем в них входят клетки ///, IV и VI слоев. По терминологии Хыобела и Визела, простые рецептивные зоны подобны сетчаточным и зонам латерального коленчатого тела, поскольку в них регистрируются «on» и «off» ответы нейронов. Однако в коре клетки обоих типов ответов расположены не концентрически, как в латеральном коленчатом теле, а вытянуты, причем отчетливо различима граница расположения возбуждающегося («on» ответы) и тормозимого («off» ответы) участков этой зоны. Эти же авторы показали, что в 18 поле простых зон нет, а, по их терминологии, 90% сложных и сверхсложных, последних :Д в 19 поле (Хьюбел и Визел, 34). Сложными были названы зоны, ответы которых на свет не могут быть предсказаны, так как в них нет разделения на возбудимый и тормозимый участки. Предполагают, что сложные зоны состоят из нескольких простых зон, конвергирующих на одну корковую клетку зоны высшего порядка.
По исследованиям же Юнга (36), в зрительной коре найдены пирамидные клетки, реагирующие не только на зрительное раздражение, но и на раздражения лабиринтных рецепторов, а также неспецифической восходящей ретикулоталамической системы. У кошки, и особенно у обезьяны, обширные
участки артельной области реагируют на раздражение вестибулярного аппарата (Юнг и сотр., 36) (рис. 49—//). При внеклеточном отведении некоторые отдельные клетки разряжаются импульсами возбуждения в ответ на все эти раздражения. Между тем известно, что раздражение лабиринтных рецепторов не дает зрительных ощущений.
Следовательно, корковые клетки, которые активируются при зрительном и вестибулярном, а также при ретикулярном раздражении, не являются сенсорными. Они должны принадлежать пирамидным вставочным или ассоциационным нейронам. Сам Юнг определенно не высказывается, с какими именно нейронами он имеет дело при каждом эксперименте. Но не может быть сомнения, что это были наиболее крупные вставочные или ассоциационные пирамидные клетки, а не сенсорные звездчатые нейроны. По-видимому, одни
|
|
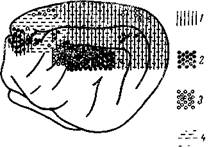
Рис. 49—II. Топографическое расположение нейронных ответов в коре большого мозга кошки на раздражение рецепторов
Дано распространение ответов: зрительных (1), слуховых на хлопки и стук (2) и вестибулярных при поляризации лабиринтов (з — ответы с коротким и 4 — ответы с длинным скрытым периодом).(По данным Юнга и сотр., Бюзе и Имберта)
и те же пирамидные нейроны принимают участие как в интеграции зрительного восприятия, так и лабиринтного. Поэтому эти нейроны должны реагировать в обоих случаях.
А пирамидные нейроны, участвующие в интеграции лабиринтного восприятия, потому, по-видимому, находятся в зрительной области, что с поворотом глаз поворачивается и голова. Следовательно, каждый раз новый поток зрительных импульсов совпадает с потоком импульсов от лабиринтных рецепторов. И потому можно считать биологически оправданной ковергенцию зрительных и лабиринтных импульсов на одних и тех же пирамидных нейронах в первичной зрительной зоне.
Но зрительная область кошки реагирует очень активно и на уколы в кожу на конечностях, в то же время не отвечая совсем на прикосновение или на движение суставов. Ответы на уколы наступают всегда с большим скрыты периодом около 70 мсек, значит через неспецифическую систему. Эта же область отвечает и на звук через ту же систему с латентным периодом 63,5 мсек. Часто наблюдается конвергенция зрительных, слуховых и соместетических (кожно-болевых) импульсов на одном и том же нейроне. И вообще все нейроны в зрительной коре, отвечающие на звук, отвечают и на уколы (Мурата и др., 107). Мы думаем, что такая взаимосвязь зрительных, слуховых и кожно-болевых раздражений в зрительной области кошки, видимо, происходит оттого, что при уколе животное поворачивает голову и фиксирует глаза на место раздражения, производит защитные движения конечностями и при этом издает звук — визжит, как это бывает при болевом раздражении. Очевидно, афферентная импульсация от всех раздражений рецепторов через неспецифическую систему направляется, между прочим, и в зрительную область, где возбудимость значительно повышается в момент фиксации глаз на раздражаемый участок кожи. Конечно, такое же взаимодействие импульсов от означенных рецепторов, должно быть, происходит и в кожной воспринимающей области, где возбудимость также должна повыситься в момент защитных движений.
Но если импульсы от сетчатки вызывают субъективное ощущение света, а из лабиринтных рецепторов не вызывают его, так это должно происходить от того, что нервные круги, возбуждаемые от сетчатки, непосредственно активируют определенный комплекс зрительных звездчатых нейронов, произ-
77
|
|
водя зрительное восприятие, а нервные импульсы, возникающие в лабиринтных рецепторах во время фиксации зрения на воспринимаемый объект и конвергирующие на пирамидных нейронах, воз буждаемых первично при зрительном восприятии, должны способствовать усиленной деятельности нервных кругов, интегрирующих сенсорные звездчатые нейроны зрения.
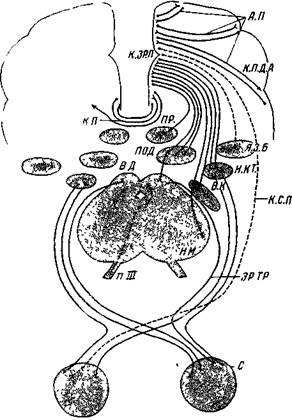
| Рис. 50. Схема эфферентных путей зрительной коры хищных Животных А.П — ассоциационные пути (короткие, средние и длинные); К ЗР. П — корковые зрительные поля, К .П . Д .А . — корковые поля других анализаторов; К .П . — комиссуральные пути (асимметричные изображения стрелкой); ПР .— претектальное ядро; ПОД. — подушка зрительного бугра, Я.З Б. — другие ядра зрительного бугра (заднее, вентральное и др.) Я.Я.Т. — наружное коленчатое тело; БД . — верхнее двухолмие; В. К. — внутренняя капсула; К С П. — кортикосетчаточный путь, Н М. — ножка мозга; ЗР.ТР. — зрительный тракт; С — сетчатка. Эфферентные пути включены в схему не полностью (Школьник-Яррос) |
Как указывалось выше, неспецифическая восходящая система также активирует корковые нейроны зрительной области. Это вполне понятно, ибо сейчас известно, что и неспецифическая таламическая афферентная система играет существенную роль вообще в активировании воспринимающих областей, которые возбуждаются посредством специфических афферентных импульсов от соответствующих рецепторов. Нужно думать, что неспецифические таламические импульсы поддерживают возбудимость пирамидных нейроров 17 поля на таком высоком уровне, что они в состоянии возбуждаться от специфических импульсов из сетчатки.
Дата добавления: 2019-07-17; просмотров: 327; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!