Активирование глиальных элементов коры.
Как указывалось выше, глия богато представлена в коре большого мозга. Между глиальными элементами, нервными элементами и сосудами существует тесная связь. Глиальные элементы заполняют все межнейрональное пространство. Часть из них так тесно связана с нервными элементами, что принято называть их сателлитами. Аксонные разветвления оканчиваются на этих глиальных сателлитах наряду с окончаниями на нервных клетках и дендритах. Поэтому можно предположить, что нейроглия играет важную роль в деятельности коры мозга. И действительно, известны такие факты, которые дают возможность приписать нейроглии самые разные функции в отношении нервной ткани мозга.
Прежде всего нейроглия должна принимать участие в обмене веществ нервных элементов. Она содержит такие ферменты, которые имеют отношение к функции дыхания (как карбоангидраза и порфирин), ибо прилегающие к нейронам глиальные элементы могут влиять на скорость их окислительных процессов (Эшби и сотр., 47). В клетках глии находят также уолинэстеразу и псевдохолинэстеразу, как в нервных клетках (Португалов, 41). Это обстоятельство указывает на возможность их участия в деятельности нервной системы. Известно активное участие глии в биосинтезе холестерина (Горизонтов, 48). А путем количественного определения рибонуклеиновой кислоты, цитохромоксидаэы и сукциноксидаэы в нервных клетках и олигодендроцитах — сателлитах ядра Дейтерса животных Хиден и Пигон (49) обнаружили, что при вестибулярной стимуляции количество означенных ферментов в олигодендроцитах уменьшается и соответствующим образом увеличивается в нервных клетках. На этом основании было сделано предположение, что афферентная импульсация первично действует на олигодендроциты, а уже эти последние влияют на нервные клетки медиаторами.
Имеются основания для предположения, что определенного типа медленная электрическая активность центральной нервной системы обусловливается нейроглией. Именно, наряду с пиковыми потенциалами клеток и аксонов, отражающими распространяющееся возбуждение, и медленными отрицательными потенциалами, отражающими местное возбуждение клеток и дендритов, в коре в ответ на раздражение возникают и значительно более медленные потенциалы. Они наблюдаются со значительно большим скрытым периодом, чем обычные медленные потенциалы нервных клеток и дендритов. Так, в ответ на вспышки света в зрительной коре удается зарегистрировать отрицательные потенциалы большой амплитуды и большой длительности (Ливанов, 50). Такие отрицательные постоянные сдвиги при периферических раздражениях наблюдали в коре и другие авторы (Касперс, 51) при выработке условных рефлексов. Очевидно, механизм генерации этих потенциалов приходит в действие и при естественных условиях возбуждения коры.
Сверхмедленные потенциалы в коре наблюдаются также и при непосредственном раздражении коры. Именно, при сильном тетаническом раздражении коры с ритмом 10—100 в сек. наступал постоянный отрицательный потенциал большой амплитуды, который нарастал при повышении частоты и силы раздражения. Во время этого потенциала дендритные отрицательные потенциалы ослабевали или сходили на нет (рис. 37 и 38) (Беритов и Ройтбак, 52).
56
|
|

Рис. 41. Хара к теристика м едленных отрицательных потенциалов
Глубокий нембуталовый наркоз; электроды на супрасильвиевой извилине, расстояние между раздражающим неактивным» отводящим электродами 0,75 мм. 1 — порог вызова дендритного потенциала (ДП); вались сделана при интенсивности раздражающего стимула 1в; 2—3 «, 3—4 < (продолжительность стимула 0,05 мсек); 4 — порог вызова медленного отрицательног о потенциала (МОП) при 10 в; в— в — на другом препарате показано возникновение МОП вследствие суммации действия подпороговых стимулов, при чем в 5 — при интенсивности стимула 7,5 в и продолжительности 0,15 мсек возникает ДП, а в б — возникает и МОП при подаче двух стимулов с интервалом 5 мсек; 7—9 — увеличение амплитуды МОП при усилении раздражения (7—5 в, 8—10 в, 9—20 в при продолжительности 0,5 мсек). Калибровка 1 м в ; 1 0—1 2 — локальность МОП. Расстояние от раздражаемого пункта до отводящего электрода соответственно — 0,75 мм, 2 мм и 3 мм (Ройтбак)
При детальном исследовании этого постоянного отрицательного потенциала Ройтбак (37) обнаружил, что он появляется при силе раздражения, в 10 раз превосходящей ту пороговую силу, при которой возникает дендритный отрицательный потенциал. При одиночных раздражениях он появляется после дендритного потенциала, т. е. со скрытым периодом не менее 20 мсек, а его продолжительность равна 0,3—0,5 сек. Но он может длиться и 3—4 сек. (рис. 41). При частых раздражениях — 10—20 в сек. амплитуда быстро достигает максимума, причем потенциал принимает постоянный неколеблющийся характер, а после прекращения раздражения потенциал затухает за 2—3 сек. Эти потенциалы отводятся на более близком расстоянии (от 2—3 мм раздражаемого участка), чем дендритные отрицательные потенциалы (рис. 42).
|
|
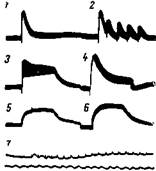
Рис. 42. Длительные отрицательные отклонения в ответ на раздражение коры большого мозга
Электроды на супрасильвиевой извилине кошки. Расстояние между раздражающими и отводящим электродами 0,75 мм. 1 — один стимул (10 в, 0,5 мсек); 2 — частота раздражения около 0,5 в сек.; з — около 10; 4— 160 в сек. Записи 5 и в произведены на другом препарате: 5 — частота раздражения 10 в сек.; в—20. Длина кадра 10 сек.; 7 — запись получена на другом препарате, частота раздражения 50 в сек. (Ройтбак)
Экспериментальный анализ показал, что эти сверхмедленные потенциалы не возникают в результате непосредственной синаптической активации дендритов, не являются результатом суммации обычных дендритных потенциалов или проявлением деятельности каких-либо нервных кругов, ибо они появляются и при глубоком нембуталовом наркозе, когда совершенно блокируется распространение возбуждения по полисинаптическим системам. Характерно, что стрихнин, который усиливает деятельность дендритов, совершенно подавляет эти медленные отрицательные потенциалы
57
|
|

Рис. 43. Действие фармакологических веществ на медленный отрицательный потенциал
Электроды на супрасильвиевой извилине, «активный» на расстоянии 0,5 мм от раздражающего. В каждом случае приводятся ответы "ч стимул (30 в, 0,05 мсек) до и после локального отравления под отводящим электродом. 1—4—действие стрихнина (0,1%); 1 — до отравления, 2 — после отравления в течение 1 мин., 3 — другой препарат, до отравления, 4 — после отравления стрихнином в течение 2_мин.; S —в — действие морфия: 5 — до отравления (электроды на другом участке извилины того же препарата), 6 — после отравления 1%-ным морфием — 5 мин.; 7—9 — действие гаммааминомасляной кислоты (ГАМН): 7 — тот же препарат, через 2 мин. после записи 6 произведено отравление того же пункта 1 % ГАМК; ответ на то же раздражение, « — другой препарат до отравления, 9 — после отравления 1 % ГАМК (Ройтбак)
(рис. 43, 1— 4 ). Также действует морфий (рис. 43, 5—6 ). Гамма-аминомасляная кислота, которая является эффективным блокирующим средством для аксодендритных синапсов, не препятствует возникновению этих медленных потенциалов (рис. 43, 7—9).
На основании приведенных фактов Ройтбак приходит к предположению, что эти постоянные потенциалы обусловлены деятельностью глиальных элементов. Как указывалось выше, глиальные элементы, в особенности олигодендроциты, находятся в прямом контакте с нервными клетками и дендритами. Олигодендроциты, например, прилегают прямо к шипикам верхушечных дендритов, а разветвления звездчатых нейронов второго типа образуют многочисленные контакты с отростками астроцитов, которые в свою очередь контактируют с нервными элементами (Шайбель и Шайбель, 53). Микроглиоциты своими многочисленными отростками, видимо, также связаны со многими нервными клетками (Микеладзе, 24). Все эти ныне хорошо известные факты поддерживают ту гипотезу, что постоянные сверхмедленные потенциалы возникают от активирования глиальных элементов.
Так как активация глиальных элементов требует приложения значительных токов, можно было подумать, что глиальные клетки раздражаются непосредственно электрическим током. Такая возможность не исключается при некоторых очень больших силах раздражения. Но Ройтбак полагает, что так как этот потенциал наступает не только в связи с усилением раздражения, то вероятнее всего, что это раздражение первично возбуждает нейроны второго и третьего слоя, а уже их аксоны активируют глиальные клетки, прежде всего олигодендроциты. А тот факт, что стрихнин и морфий устраняют длительные отрицательные потенциалы, не ослабляя дендритные потенциалы, даже благоприятствуют им, должно быть, зависит от того, что эти вещества при некоторой высокой концентрации парализуют глиальные клетки — сателлиты нейронов.
Медленные потенциалы, отводимые от поверхности коры, в значительной мере зависят, по этой гипотезе, от активации дендритов под влиянием глиальных элементов. Возникающие в дендритах потенциалы создают токи, которые, согласно дендритной гипотезе торможения, электротонически распространяясь по верхушечным дендритам, должны производить анэлектротоническую блокаду соматических синапсов, т. е. торможение пирамидных
58
|
|

Рие. 44. Изменение первичного ответа во время медленного отрицательного потенциала (МОП), вызываемого раздражением коры кошки
Отводящий микроэлектрод и раздражающие электроды находятся на задней сигмоиднойизвялине;|расстояние между электродами 0,5 мм. 1— положительный первичный ответ от поверхности коры на раздражение кожи противоположной передней лапы; 2 —эффект того же раздражения во врзмя МОП через 100 мсек после 'однократного раздражения коры (0,2 мсек, 30 в); 3—в — после углубления микроэлектрода на глубину 1600 мк; з и s — отрицательные первичные ответы от одного раздражения кожи, а 4 и в — во время МОП, вызванного раздражением коры; 7— отводится^поверхность коры макроэлектродом. Усилитель с болыдэй постоянной времени. При .том же раздражзнии коры регистрируется сначала обычный дендритный потенциал, а затем МОП (Ройтбак)
клеток. Что пирамидные клетки действительно тормозятся, это хорошо видно из рис. 44. При одновременном отведении потенциалов от поверхности коры и из глубины ее наблюдается исчезновение спонтанных разрядов и ослабление первичных потенциалов пирамидных клеток одновременно с длительным отрицательным потенциалом от поверхности коры (Ройтбак, 54).
Вышеизложенное представление о нейроглиальном происхождении медленных отрицательных потенциалов поверхности коры находит подтверждение в последних исследованиях, произведенных с помощью микроэлектродов, вводимых в отдельные глиальные клетки, располагающиеся между нервными волокнами в зрительном нерве амфибий. При электрическом раздражении нерва или при адекватном раздражении сетчатки в глиальных клетках развивается длительная деполяризация мембраны. В ответ на однократное раздражение она длится несколько секунд. При ритмических раздражениях сдвиги мембранного потенциала суммируются; записи подобны записям медленных корковых потенциалов (Каффлер, Никколс, 55, 56). Каффлер на основании ряда фактов считает вероятным, что причиной деполяризации глиальной мембраны является выход ионов калия из нервных волокон при их возбуждении и накоплении ионов калия в межклеточных щелях; но можно думать, что в коре и вообще в центральной нервной системе деполяризация глиальной мембраны происходит в результате выделения медиатора из контактирующих с глиальными клетками окончаний нервных волокон (Ройтбак, 54).
Исходя из вышеизложенного, надо полагать, что н ейрогл ия коры долж на играт ь существенную рол ь в целостной деятел ь ности головного мозга.
Нейроглия настолько разнообразна и настолько тесно связана с нервной системой, настолько она мощно развита, намного превосходя количественно нервную систему, что видный нейрофизиолог Галамбос (57) счел возможным приписать ей самую высшую функцию мозга — сознательную деятельность.
59
ЛИТЕРАТУРА
1. Поляков Г. И. и СаркисовС. А. В кн. «Цитоархитектоннка коры большого мозга человека». Медгиз, 1949, 102.
2. Школьник-ЯрросЕ. Г. Проблемы физиол. оптики, сб. II. 1955, 102.
3. О' L е а гу J. H. J. Compar. Neurol., 1941, 25, 131.
4. LoreatedeN6R. J. Psychol. Neurol., 1933, 45, 381; 1934, 46, 113; In: Fulton J. F. «Physiology of the nervous system», 1944, 274.
5. 0' L е а г у J. H. а. В i sh о p G. H. J. Gompar. Neurol., 1938, 68, 423.
6. П о л я к о в Г. И. Архив анат., гистол. и эмбриол., 1953, 30, 48.
7. Jasper H.,Naquet P. a. KingE. EEG and Clin. NeurophysioL, 1955, 7, 99.
8. L i С. L. J. NeurophysioL, 1959, 22, 436.
9. Поляков Г. И. Тезисы докл. VI Всесоюзн. съезда анат., гистол. и эмбриол., 1958, 78.
10. С h a n g Н. Т. Cold Spr. Harb. Biol. Sympos., 1952,17, 189.
11. Э н т и н Т. И. Архив анат., гистол. и эмбриол., 1954, 31, 218.
12. С а р к и с о в С. А. Некоторые особенности структуры нейрональных связей в коре головного мозга. М., Медгиз, 1948.
13. Hernandez-Peon R., Scherrer H. a. JouvetM. Science, 1956, 123, 331; Fed. Proc., 1956, 15, 91.
14. J u n g R. Annales de la Facultad de Medicin, Montevideo, 1959, 44, N 3-4, 323; Structure a, function of the cerebral cortex. Proc. of the Second Intern. Meeting of Neurologists, Amsterdam, 1953j 204.
15. P e n f i e 1 d W. Amer. Res. Nerv. Dis. Proc., 1952, 30, 519; The excitable cortex in conscious man. Liverpool, 1958.
16. Школьник-ЯрросЕ. Г. Тезисы докл. VI Всесоюзн. съезда анат., гистол. и эмбриол., 1958, 436.
17. Б е р ит ов И. С. Общая физиология мышечной и нервной системы, т. II. Спинной мозг и ствол головного мозга. М., «Медицина», 1966.
18. П о л я к о в Г. И. Журн. высш. нервн. деят., 1956, 8, 469.
19. С a j а 1 8. R a m о n у. Studies on the cerebral cortex. Lloyd-Luke, 1955.
20. В е р и т а ш в и л и И. С. Труды Ин-та физиол. АН ГССР, 1956, 10, 3.
21. Ж у к о в а Г. П. Архив анат., гистол. и эмбриол., 1953, 30, 1.
22. С у х а н о в С. A. La cellula, 1898, 14, 2.
23. Р и г р и г a D. P. Internal. Rev. Neurobiol., 1959,1, 47. В кн. «Механизмы целого мозга». М., ИЛ, 1963, 10.
24. М и к е л а д з е А. Л. Электронная микроскопия центральной нервной системы. Докт. дисс. Тбилиси, 1966.
25. Jasper H.,Stefanis С. EEG and Clin. Neurophysiol., 1965, 18, 541.
26. Li C. L. J. Cell. a. Compar. Physiol., 1961, 58, 155; 1962, 60, 1.
27. С r e u t z f e 1 d t 0. D., W a t a n a b e S., Lux H. D. EEG and Clin. Neurophysiol., 1966, 20, 19.
28. Phillips C. G. Quart. J. Exp. Physiol., 1959, 44, N 1, 1.
29. E с с 1 e s J. The physiology of the nerve cells. Baltimore, 1957 [Э к к л с Дж. «Физиология нервных клеток». М., ИЛ, 1959].
30. Н i I d W., Т a s a k i J. J. NeurophysioL, 1962, 25, N 2, 277.
31. Зурабашвили А. Д. Синапсы и обратимые изменения нервных клеток. М., 1951.
32. Maximow А. [Максимов А. ]А text-book of Histology. 1931.
33. Р u r p u r a D. P. Ann. N. Y. Acad. Sci., 1961, 92, 840; 1961, 94, 604.
34. P о и т б а к А. И. Труды Ин-та физиол. АН ГССР, 1953, 9, 97; В кн. «Современные проблемы электрофизиологических исследований нервной системы». М., 1964, 171.
35. В u r n s В. D. J. Physiol., 1954, 152, 427.
36. P о и т б а к А. И. Биоэлектрические явления в коре больших полушарий. Тбилиси, изд-во АН ГССР, 1955.
37. Р о и т б а к А. И. В кн. «Современные проблемы деятельности и строения ЦНС». Тбилиси, изд-во «Мецниереба», 1965, 67.
38. Е с с 1 е s J. C. EEG and Clin. NeurophysioL, 1951, 3, 449.
39. С 1 a r e М. Н. а. В i s h о p G. H. EEG and Clin. NeurophysioL, 1955, 7, 85v
40. A d r i a n E. D. J. Physiol., 1936, 88, 127.
41. П о р т у г а л о в В. В. Тезисы Докл. VI Всесоюзн. съезда анат., гистол. и эмбриол. Харьков, 1958, 84; Гистохимические методы в нормальной и патологической морфологии. М., 1958, 28.
42. Э к к л с Дж. Физиология синапсов. М., «Мир», 1966.
43. Беритов И. С. Нервные механизмы поведения высших позвоночных животных. Изд-во АН СССР, 1961.
44. С h a n g Н. Т. J. NeurophysioL, 1951, 14, 1, 23.
45. W о о 1 s е у С. N. а. С h a n g H. T. In: «The frontal lobes». Baltimore, 1948, 146.
46. M a g о u n H. W. The waking brain. Ch. C. Thomas Publishers. USA, 1958. [М э г у а Бодрствующий мозг. Изд. 2. М., «Мир», 1966].
47. A s h Ь у W., G о z z е 1 i R., S с h u s t e r А. Е. Цит. по: Н у d е и Н. «The neuron». 1967.
60
48. Г о р и з о н т о в П. Д. Значение головного мозга в холестериновом обмене. М., 1940.
49. Н у d е n Н.. Р i g 6 n A. J. Neurochem., 1960, 6, 57.
50. ЛивановМ. Н. В кн. «Электроэнцефалографические исследования высшей нервной деятельности». Изд-во АН СССР, 1962. 174.
51. Gaspers H. Pfl. Arch. ges. Physiol., 1959, 269, 157.
52. Б е р и т о в И. С., Р о и т б а к А. И. Журн. высш. нервн. деят., 1955, 5, 173.
53. S с h е i b е 1 М. Е. a. S с h е i Ь е 1 А. В. Progress in brain research, 1964, 9, 6.
54. Р о и т б а к А. И. Международная конференция, посвященная столетию выхода в свет труда И. М. Сеченова «Рефлексы головного мозга». Изд-во «Наука», 1965, 186.
55. N i с h о 11 s J. G. а. К u f f 1 е г S. W. J. Neurophysiol., 1964, 27, 645.
56. О г k a n d R., N i с h о 11 s J. а. К и f f 1 е г S. J. Neurophysiol., 1966, 29, 769 а. 788.
57. G a 1 a m b о s R. Proc. Nat. Acad. Sci. (Wash.), 1961, 47, 129.
58. Р и г р и r a D. P., G i r a d о M., G г и n d f e s t H. Science, 1957, 125, 1200.
59. О к у д ж а в а В. М. Активность верхушечных дендритов в коре больших полушарий. Тбилиси. Изд-во АН ГССР, 1963.
60. Р о и т б а к А. И. В кн. «Центральные и периферические механизмы нервной деятельности». Ереван. Изд-во АН Арм. ССР, 1966, 365.
61. G о 1 d г i n g S., 0* L е а г у J. L. Fed. Proc., 1960, 19, 612.
62. J a s р е r H. In: «Inhibition in the nervous system and GABA». PergamonPress, Oxford — N. Y.— London — Paris, 1960, 544.
63. Р о и т б а к А. И. Влияние стрихнина на дендритные мембраны. 1967. Не опубликовано.
64. Е с с 1 е s J. С. Ergebn. Physiol., 1961, 51, 300; Е с с 1 е s J. С., К о s t у u k P. а. S ch m i d t R. J. Physiol., 1962, 161, 237; 1962, 162, 138.
Дата добавления: 2019-07-17; просмотров: 350; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!