Эндокринная часть половых желез
Яичко у мужчин и яичник у женщин, помимо половых клеток, вырабатывают и выделяют в кровь половые гормоны, под влиянием которых формируются вторичные половые признаки.
Эндокринной функцией в яичке обладает интерстиций, представленный железистыми клетками – интерстициальными эндокриноцитами (клетки Лейдига), располагающимися в рыхлой соединительной ткани между извитыми семенными канальцами, кровеносными и лимфатическими капиллярами. У взрослого человека эндокриноциты локализуются либо в виде скоплений возле сосудов (собственно интерстициальные), либо рядом с канальцами (перитубулярные эндокриноциты).
Интерстициальные эндокриноциты яичка выделяют основной мужской половой гормон – тестостерон, а также небольшое количество окситоцина и производных пропиомеланокортина. Эти гормоны регулируют течение сперматогенеза, а также развитие и функцию добавочных желез половой системы, обеспечивают развитие вторичных половых признаков, определяют либидо (половое влечение) и половое поведение. Окситоцин контролирует сократительную активность миоидных перитубулярных клеток извитых семенных канальцев.
В последние годы показано, что простата также обладает гормонпродуцирующей функцией, эндокринные клетки которой вырабатывают серотонин, соматостатин, пептиды кальцитонинового ряда, оказывающие влияние на секреторную активность желез органа и сократительную деятельность гладко‑мышечных клеток стромы, а также подвижность сперматозоидов.
В яичнике вырабатываются половые гормоны: эстроген, гонадокринин и прогестерон. Местом образования эстрогена (фолликулина) и гонадокринина являются зернистый слой созревающих фолликулов, а также клетки интерстиция яичника. Эстроген стимулирует, а гонадокринин угнетает рост и развитие половых клеток.
Под влиянием фолликулостимулирующего и лютеинизирующего гормонов гипофиза осуществляются рост фолликула и активация интерстициальных клеток. Лютеинизирующий гормон вызывает овуляцию и образование желтого тела – своеобразного, обладающего эндокринной функцией органа, клетки которого вырабатывают гормон яичника прогестерон. Прогестерон подготавливает слизистую оболочку матки к восприятию оплодотворенной яйцеклетки, задерживает рост новых фолликулов, а также способствует нормальному протеканию беременности. Эти же клетки продуцируют также эстрогены и в небольшом количестве андрогены и окситоцин, а при беременности – полипептидный гормон релаксин, который подготавливает родовые пути к родам.
Эндокринная часть поджелудочной железы
В поджелудочной железе, помимо внешней секреторной (экзокринной) части, имеется внутрисекреторная (эндокринная) часть, представленная панкреатическими островками (островки Лангерганса – Соболева), общее количество которых у взрослого достигает 1,5–2 млн, составляя 1–2 % от объема органа. Это овальные или округлые образования величиной 0,1–0,8 мм, располагающиеся по всей толще железы; больше их в хвостовом отделе. Островки состоят из эпителиальных клеток (инсулоцитов), окруженных соединительной тканью, содержащей густую сеть фенестрированных кровеносных капилляров и нервных волокон. Выделяют пять основных типов инсулоцитов (А, В, D, D1и РР). А‑клетки, составляющие 20–25 % от общего числа инсулоцитов, располагаются преимущественно по периферии островков. Они секретируют гормон глюкагон, повышающий уровень глюкозы в крови вследствие расщепления гликогена и липидов. В‑клетки (60–70 % от всех инсулоцитов) образуют центральную часть островков и вырабатывают гормон инсулин, который стимулирует синтез гликогена и поглощение глюкозы клетками различных тканей, вызывая снижение ее уровня в крови. D‑клетки (5–10 % от всех инсулоцитов) располагаются на периферии островков, выделяют в кровь гормон соматостатин, оказывающий многообразное действие, в частности угнетает секрецию А– и В‑клеток, а также панкреатоцитов. D‑клетки рассматриваются как разновидность предыдущих и вырабатывают вазоактивный интестинальный пептид (ВИП) – гормон, вызывающий снижение давления крови, а также усиление выработки панкреатического сока. РР‑клетки, составляющие 2–5 % от всех инсулоцитов, обычно располагаются по периферии островков, но могут встречаться и в экзокринной части железы. Они секретируют панкреатический полипептид – гормон, угнетающий активность панкреатоцитов.
Поражение клеток островков, например в результате аутоиммунного процесса, приводящее к их разрушению, вызывает развитие сахарного диабета – распространенного эндокринного заболевания, ведущую роль в течение которого играет недостаточность инсулина.
Островки Лангерганса – Соболева относятся к гастроэнтеропанкреатической системе эндокринных клеток, являющихся частью диффузной эндокринной системы (ДЭС). ДЭС образована эндокриноцитами, рассеянными по различным органам и тканям, где они располагаются поодиночке или мелкими группами. Значительное их число находится в слизистых оболочках различных органов и связанных с ними железах. Они особенно многочисленны в пищеварительном тракте, но наблюдаются и в дыхательной, мочевой, половых, сердечно‑сосудистой системах, слюнных железах, органах чувств и т. д. Предполагают, что эти клетки участвуют в анализе химического состава пищи, воздуха, мочи, крови и т. п. и отвечают на его изменения выделением гормонов и паракринных факторов. Общее количество клеток ДЭС в несколько раз превышает число клеток эндокринных органов, а их секреторные продукты оказывают как местное (паракринное), так и дистантное (эндокринное) влияние на моторику гладко‑мышечных клеток в стенках различных органов, секрецию экзо– и эндокринных желез и т. д. Учение о ДЭС – одно из самых перспективных интенсивно развивающихся научных направлений, имеющее не только теоретическое, но и большое практическое значение для медицины.
Тесты и вопросы для самоконтроля
Выберите один или несколько правильных ответов:
1. К центральным органам эндокринной системы относятся:
а) надпочечники;
б) гипоталамус;
в) шишковидная железа;
г) околощитовидные железы;
д) гипофиз.
2. Шишковидная железа выполняет следующие функции:
а) регулирует тонус скелетной мускулатуры;
б) регулирует циркадные ритмы организма;
в) контролирует половое созревание организма;
г) повышает кровяное давление.
3. Нейросекреторные клетки переднего гипоталамуса вырабатывают гормоны:
а) вазопрессин;
б) альдостерон;
в) окситоцин;
г) адренокортикотропный гормон;
д) кальцитонин.
4. Передняя доля гипофиза (аденогипофиз) включает:
а) воронку, дистальную, бугорную части;
б) дистальную, бугорную, промежуточную части;
в) дистальную, бугорную, нервную части;
г) воронку, бугорную и нервную части.
5. Ацидофильные клетки аденогипофиза продуцируют:
а) соматотропин;
б) пролактин;
в) тиреотропный гормон;
г) липотропный гормон;
д) лютеинизирующий гормон.
6. Базофильные клетки аденогипофиза секретируют:
а) адренокортикотропный гормон;
б) рилизинг‑факторы;
в) тиреотропный гормон;
г) фолликулостимулирующий гормон;
д) лютеинизирующий гормон;
е) пролактин.
7. Гормонами щитовидной железы являются:
а) пролактин;
б) трийодтиронин;
в) тетрайодтиронин;
г) соматотропин;
д) кальцитонин.
8. Околощитовидные железы секретируют гормоны:
а) кальцитонин;
б) тиреотропный гормон;
в) паратгормон;
г) тетрайодтиронин.
9. Гормонами коркового вещества надпочечника являются:
а) минералкортикоиды;
б) глюкокортикоиды;
в) кальцитонин;
г) половые стероиды;
д) пролактин.
10. Мозговое вещество надпочечника продуцирует:
а) альдостерон;
б) адреналин;
в) норадреналин;
г) андрогены;
д) кальцитонин.
Ответы
1: б, в, д.
2: б, в.
3: а, в.
4: б.
5: а, б.
6: а, в, г, д.
7: б, в, д.
8: в.
9: а, б, г.
10: б, в.
Глава 14
Пищеварительная система
Пищеварительная система включает пищеварительный тракт и крупные железы: слюнные, печень и поджелудочную железу. Функции:
• пищеварительная – механическая и химическая обработка пищи, всасывание питательных веществ и удаление непереваренных веществ;
• экскреторная – выведение продуктов обмена и других вредных веществ через стенку пищеварительного тракта;
• иммунная – захват антигенов с последующим развитием иммунной реакции;
• эндокринная – выработка гормонов (инсулин, глюкагон, ВИП, гастрин и др.).
Общим является наличие трех оболочек: слизистой, мышечной и серозной или адвентициальной. Слизистая оболочка (tunica mucosa):
1) эпителий представлен однослойным призматическим в желудке, тонкой и толстой кишках, многослойным неороговевающим в полости рта, пищеводе, анальной части прямой кишки, а также бокаловидными и эндокринными клетками. Занимает пограничное положение с внешней средой, образуя барьер с избирательной проницаемостью, обеспечивающий переваривание и всасывание питательных веществ. На поверхность эпителия выделяют секреты железы, продуцирующие и выделяющие слизь;
2) собственная пластинка слизистой оболочки располагается под эпителием и представлена рыхлой волокнистой соединительной тканью, в которой расположены нервные волокна, диффузная лимфоидная ткань; в пищеводе и желудке располагаются также простые железы, в миндалинах – крипты (углубления эпителия). В полости рта собственная пластинка образует сосочки, в тонкой кишке – ворсинки;
3) мышечная пластинка слизистой оболочки образована гладко‑мышечными клетками, формирующими внутренний циркулярный и наружный продольный слои, сокращение которых способствует изменению ее рельефа;
4) подслизистая основа обеспечивает подвижность слизистой оболочки; образована рыхлой волокнистой соединительной тканью, содержащей сплетения кровеносных и лимфатических сосудов, подслизистое нервное сплетение (мейсснеровское сплетение), лимфоидную ткань, концевые отделы желез (в пищеводе и двенадцатиперстной кишке).
Мышечная оболочка (tunica muscularis) на основном протяжении пищеварительной трубки представлена гладко‑мышечной тканью, в переднем отделе (полость рта, глотка, верхняя треть пищевода, заднепроходный канал) – поперечнополосатой (скелетной) мышечной тканью. Состоит из двух слоев – наружного продольного и внутреннего циркулярного, между которыми располагается прослойка рыхлой соединительной ткани, содержащая сосудистое и межмышечное нервное (ауэрбаховское сплетение) сплетения. Сокращения мышечной оболочки обеспечивают перемещение и продвижение содержимого пищеварительной трубки.
Наружная оболочка (tunica serosa, s. adventitia) серозная или адвентициальная:
1) серозный висцеральный листок брюшины – состоит из слоя соединительной ткани и мезотелия. Покрывает большую часть поддиафрагмального отдела пищеварительного тракта;
2) адвентициальная оболочка образована рыхлой волокнистой соединительной тканью; составляет наружную оболочку органов наддиафрагмального отдела.
Полость рта
Полость рта включает губы, щеки, зубы, десны, твердое и мягкое небо, язык, миндалины.
В губах различают кожную часть (покрыта многослойным плоским ороговевающим эпителием, имеются сальные, потовые железы, волосы), промежуточную часть (покрыта многослойным плоским ороговеваюшим эпителием, собственная пластинка слизистой оболочки богато васкуляризована и иннервирована) и слизистую часть (покрыта многослойным плоским неороговевающим эпителием, собственная пластинка слизистой оболочки образует высокие сосочки, имеются губные слизистые железы).
Щека. Внутренняя поверхность щеки выстлана многослойным плоским неороговевающим эпителием, собственная пластинка представлена плотной фиброэластической тканью, образует высокие сосочки. Хорошо выражена подслизистая основа (эластическая и жировая ткани, мелкие слизистые железы). Различают верхнечелюстную, нижнечелюстную и промежуточную зоны щеки.
Небо (мягкое и твердое). Мягкое небо – это складка слизистой оболочки с фиброзно‑мышечной основой, отделяющая полость рта от глотки. Выделяют переднюю ротоглоточную поверхность, выстланную многослойным плоским неороговевающим эпителием; собственная пластинка образует высокие сосочки, подслизистая основа содержит малые слюнные железы, жировую ткань; заднюю носоглоточную поверхность, выстланную многорядным призматическим реснитчатым эпителием; в собственной пластинке встречаются лимфоидные узелки.
Твердое небо покрыто многослойным плоским, частично ороговевающим эпителием; подслизистая основа отсутствует. Сращено с надкостницей небных отростков верхних челюстей.
Десна. Это слизистая оболочка, окружающая зуб, выстлана многослойным плоским, частично ороговевающим эпителием; в собственной пластинке слизистой оболочки много капилляров и нервных окончаний.
Язык. Образован поперечнополосатой скелетной мышечной тканью. Пучки располагаются в трех взаимно перпендикулярных плоскостях. Нижняя поверхность языка покрыта многослойным плоским неороговевающим эпителием, хорошо выражена подслизистая основа.
Анатомически выделяют тело (передние 2/3) и корень языка (задняя треть), разделяющиеся V‑образной пограничной бороздой. На верхней части языка слизистая оболочка образует характерные выпячивания – сосочки, покрытые многослойным плоским, частично ороговевающим эпителием, собственная пластинка сращена с мышцами языка. Различают 4 типа сосочков: нитевидные, грибовидные, желобовидные, листовидные (у детей). Между сосочками располагаются щели и ровики, в которых открываются протоки серозных слюнных желез.
1. Нитевидные сосочки самые многочисленные, имеют заостренную, коническую форму, их многослойный плоский ороговевающий эпителий образует характерный белый налет языка.
2. Листовидные сосочки развиты в раннем детском возрасте на боковых поверхностях языка, у взрослых рудиментарны. На боковой поверхности сосочков располагаются вкусовые луковицы.
3. Грибовидные сосочки имеют узкое основание и шляпку в виде гриба. Сквозь тонкий многослойный плоский неороговеваюший эпителий просвечивают капилляры, расположенные в собственной пластинке. Сосочки красного цвета, на боковых поверхностях их расположены вкусовые луковицы.
4. Желобовидные (окруженные валиком) сосочки – самые крупные, располагаются по пограничной борозде языка в количестве 6–15. Окружены валиком (утолщение слизистой оболочки), в ровик открываются серозные слюнные железы. На боковой поверхности имеются многочисленные вкусовые луковицы.
Лимфоэпителиальное глоточное кольцо Пирогова – Вальдейера включает миндалины – лимфоэпителиальные органы, выполняющие защитную функцию (лимфоциты, макрофаги, иммуноглобулины); наибольшего развития достигают в детском возрасте, но подвергаются инволюции после полового созревания.
Язычная миндалина складчатая, складки обусловлены скоплением лимфоидных фолликулов в собственной пластинке слизистой оболочки. Фолликулы имеют центры размножения, между которыми располагается диффузная лимфоидная ткань. Эпителий многослойный плоский неороговевающий, погружаясь в лимфоидную ткань, образует крипты, в которые открываются протоки слизистых желез.
Небная миндалина располагается между небными дужками, окружена плотной соединительно‑тканной капсулой. Эпителий многослойный плоский неороговеваюший, образует углубления – крипты, инфильтрирован лимфоцитами, макрофагами и плазматическими клетками, а также дендритными антиген‑представляющими клетками. В собственной пластинке располагаются лимфоидные узелки с герминативными центрами, между ними – межузелковая диффузная лимфоидная ткань, а также соединительная ткань.
Глоточная миндалина расположена на задней поверхности глотки. Эпителий многорядный призматический реснитчатый, инфильтрирован лимфоцитами. В собственной пластинке слизистой оболочки находятся концевые отделы смешанных белково‑слизистых желез.
Трубные миндалины – это скопления лимфоидной ткани в области глоточных отверстий слуховых труб. Эпителий многорядный призматический реснитчатый. Строение сходно со строением глоточной миндалины.
Зубы. Это твердые органы полости рта, обеспечивающие пережевывание пищи. Зуб состоит из коронки, выступающей над поверхностью десен, и одного или нескольких корней, погруженных в альвеолу челюсти и прикрепленных к ней периодонтальной связкой, или периодонтом.
Основу зуба составляет твердая обызвествленная ткань – дентин, который снаружи покрыт двумя другими обызвествленными тканями: в области корня – цементом, а в области коронки – эмалью. Эмаль и цемент контактируют в шейке зуба, к которой прикреплен эпителий десны.
Внутри зуба, в пульпарной камере, находится пульпа – рыхлая соединительная ткань, содержащая сосуды и нервы, проникающие в зуб через верхушечное отверстие их корня.
Эмаль (enamelum) покрывает коронку зуба, формируя весьма прочный и устойчивый против стирания слой. Толщина слоя эмали в различных отделах коронки постоянных зубов неодинакова и колеблется от 1,7–2,5 мм на уровне жевательных бугорков моляров до 0,01 мм в области шейки зуба. Молочные зубы имеют слой эмали, не превышающий 0,8–1,0 мм.
Зрелая эмаль является самой твердой тканью в теле человека. По твердости ее нередко сравнивают с кварцем. Наибольшей твердостью обладают поверхностные слои эмали. По направлению к дентино‑эмалевому соединению она постепенно снижается. Прочность эмали самая высокая у режущего края и уменьшается к пришеечной части коронки зуба.
Твердость эмали связана в основном с тем, что эмаль на 96–99 % состоит из неорганической основы (соли) и лишь на 1–4 % из органической (белки и вода). Неорганическую основу представляет фосфат кальция в форме гидроксиапатитных кристаллов, что делает ее схожей с другими минерализованными тканями тела (кость, дентин, цемент). Уникальность эмали состоит в следующем:
• она происходит из эктодермы;
• ее органическая основа (матрица) неколлагенна по своей природе;
• кристаллы эмалевого апатита намного больше кристаллов других минерализованных тканей;
• в зрелой эмали нет клеток. На определенном этапе амелогенеза клетки резорбируются, тогда как новые не появляются. В связи с этим рост зрелой эмали отсутствует.
Эмаль исключительно твердая, что позволяет ей успешно противостоять усилиям жевания и другим механическим нагрузкам. Однако она хрупкая, и если бы не подлежащий упругий дентин, то эмаль постоянно растрескивалась бы при значительной нагрузке.
Несмотря на такую твердость, зрелая эмаль обладает определенной проницаемостью для веществ, находящихся в слюне или пище.
Цвет эмали может значительно меняться от желтого до различных оттенков серо‑белого или серого. Это зависит от прозрачности ткани: чем она прозрачнее, тем больше просвечивает дентин желтого цвета. Оттенки голубого наиболее заметны у режущего края, где нет подлежащего дентина. Прозрачность эмали, вероятно, связана с высокой степенью минерализации и гомогенностью ткани, на нее не влияет толщина эмали.
Эмаль условно называют тканью, поскольку она не содержит клеток, а является производной эпителия, который ее секретирует и минерализует.
Структурно‑функциональные особенности эмали обусловлены определенными количественными взаимоотношениями органических и неорганических веществ (матриц) эмали.
Органическая матрица эмали. В отличие от органических матриц других минерализированных тканей, формирующихся на основе коллагенов, органическая матрица эмали состоит из неколлагеновых протеинов с общим названием «эмалевые протеины» (белки). Эмалевые протеины являются гликозилированными полипептидами, синтезированными и выделенными (посредством мерокринного механизма) амелобластами. Выделяют два различных класса эмалевых протеинов – энамелины и амелогенины.
Энамелин является кислым гликопротеином с высокой мол. массой – 72 000. Амелогенин – это гидрофильный, богатый пролином гликопротеин с мол. массой 30 000.
Энамелины характеризуются также высоким содержанием глицина, серина (фосфосерина), аспарагиновой и глутаминовой кислот (последняя, вероятно, в форме γ‑карбоксиглутаминовой кислоты, имеющей тесное сродство к кальцию). Амелогенины в дополнение к пролину обогащены лейцином, гистидином и глутаминовой кислотой.
В органической матрице эмали также присутствуют гликозаминогликаны, протеогликаны и различные классы липидов. Эти органические вещества выполняют определенную роль в кальцификации эмалевых протеинов.
Неорганическая матрица эмали. Неорганическое вещество эмали представлено [Боровский Е. В., 1973] следующим составом (в процентах): гидроксиапатит – 75,04; карбонатапатит – 12,06; хлорапатит – 4,39; фторапатит – 0,65; карбонат кальция – 1,33; карбонат магния – 1,62.
В составе соединений кальция 37 %, а фосфора 17 %.
Таким образом, как и в кости, дентине и цементе, основным минералом эмали является кальция фосфат в форме кристаллической гидроксиапатитной решетки (Са10(РО4)6ОН2). В отличие от других кальцинированных тканей кристаллы апатита в виде пластинок гексагональной формы в эмали исключительно велики. Их длина достигает 500–600 нм (в среднем около 200 нм). В кости и дентине кристаллы апатита небольшие, тонкие, игольчатые. При образовании кристаллического гидроксиапатита не возникает аморфного фосфата кальция, как в кости или дентине.
Кристаллы апатита эмали, помимо кальция, содержат фосфаты и гидроксильные остатки, а также натрий, магний, карбонаты, имеющиеся в достаточном количестве в эмали около дентино‑эмалевого соединения. Концентрация этих веществ резко уменьшается к поверхности эмали. В поверхностных слоях эмали в большей концентрации обнаружены такие элементы, как фтор, свинец, цинк, содержание которых уменьшается в глубоких слоях зрелой эмали. Этот ионный градиент имеет определенное функциональное значение. Например, фтор, свинец и цинк, включенные в эмалевый гидроксиапатит, придают его кристаллам исключительную прочность и сопротивляемость кислотам. Такая эмаль имеет высокую сопротивляемость кариесу, поскольку проницаемость на ее поверхности значительно снижена.
Ионы, связанные с кристаллами гидроксиапатита эмали, передают ее глубоким слоям больше воды и растворимых кислот. В связи с этим такая эмаль менее подвержена болезням и обладает высокой сопротивляемостью к раскалыванию вдоль дентино‑эмалевого соединения.
В эмали находится также незначительное количество алюминия, бария, меди, молибдена, стронция, серы, олова и титана.
Поверхностный слой эмали отличается от остальной части своими физико‑химическими характеристиками. Поверхность эмали непроходима для рентгеновского излучения, тверже и менее проницаема по сравнению с более глубокими слоями эмали, она содержит в 5–10 раз больше фтора и более высокую концентрацию углеводородов.
В эмали нет клеток, и она не способна к регенерации, однако в ней постоянно происходит обмен веществ (в основном ионов), поступающих в нее как со стороны подлежащих зубных тканей (дентин, пульпа), так и из слюны. Поступление ионов одновременно сопровождается их удалением из эмали (деминерализация), что увеличивает или уменьшает проницаемость эмали.
Сдвиг процесса в ту или иную сторону зависит от ряда факторов, в том числе от содержания макро– и микроэлементов в слюне, рН в полости рта и на поверхности зуба.
Степень проницаемости эмали неодинакова в различные периоды развития зуба. Она снижается с возрастом, поэтому эмаль постоянного зуба молодого человека более проницаема, чем эмаль зуба пожилого человека.
Эмалевые призмы. Структурно эмаль состоит из эмалевых призм и склеивающего их межпризматического вещества.
Призмы, число которых составляет несколько миллионов в каждом зубе, представляют собой тонкие удлиненные образования, проходящие через всю толщу эмали. Толщина призм колеблется от 3 до 5 мкм, а длина различна в разных отделах коронки зуба, как правило, она больше толщины слоя эмали. Вследствие этого эмалевые призмы, собранные в пучки, образуют волнообразные изгибы по своему ходу, изогнутые в виде буквы S. В связи с этим на шлифах зуба невозможно проследить ход каждой отдельной призмы от дентино‑эмалевой границы до поверхности зуба.
Эмалевые призмы располагаются в радиальном направлении, т. е. под прямым углом к дентино‑эмалевому соединению. В области жевательных бугорков или режущего края зубов они лежат параллельно длинной оси зуба, а на боковых поверхностях коронки постепенно перемещаются в плоскость, перпендикулярную к длинной оси зуба, или даже несколько уклоняются от нее в сторону верхушки корня.
Линии Гунтера – Шрегера. Вследствие того что эмалевые призмы имеют по своему ходу S‑образную изогнутость, на продольном шлифе зуба не удается разрезать каждую эмалевую призму строго продольно на всем ее протяжении: одни участки призм оказываются сошлифованными в продольном направлении, а их продолжения в поперечном или косом. Правильное чередование поперечных (диазоны) и продольных (паразоны) шлифов пучков эмалевых призм при их изучении в отраженном свете предстает как чередование темных и светлых полос, пересекающих в радиальном направлении всю толщу эмали и называемых полосами Гунтера – Шрегера; они хорошо заметны даже при малом увеличении. Начинаясь от дентино‑эмалевого соединения, эти полосы в виде темных и светлых дуг идут кнаружи, заканчиваясь недалеко от наружной поверхности эмали.
Полоски Ретциуса. В эмали часто бывают видны линии, или полоски, Ретциуса, пересекающие полосы Гунтера – Шрегера под острым углом. Как правило, они бывают окрашены в желтовато‑коричневый цвет, усиливающийся с возрастом.
Наиболее многочисленные и вместе с тем наиболее короткие линии Ретциуса имеются в эмали боковых поверхностей коронки зуба. Начинаясь от дентино‑эмалевой границы, они косо пересекают всю толщу эмали и заканчиваются на ее поверхности бороздами, отделенными друг от друга высокими валиками. Эти валики, высотой 2–4 мкм и шириной 30–150 мкм, располагаются горизонтально параллельными рядами, опоясывая всю окружность зуба. Они носят название «перикиматии» и особенно отчетливо видны в пришеечной области. Перикиматии исчезают с возрастом в связи со стиранием поверхности эмали. На молочных зубах они выражены значительно слабее, чем на постоянных.
На дне бороздок имеются многочисленные мелкие вдавления (ямки) на поверхности эмали глубиной 0,5–3,0 мкм, которые появляются в процессе развития и соответствуют расположению отростков Томса амелобластов на завершающих стадиях секреции эмали.
По направлению к жевательной поверхности зуба линии Ретциуса делаются более длинными. Некоторые из них, начинаясь у дентино‑эмалевой границы: на боковой поверхности зуба, дугообразно огибают область жевательного бугорка и заканчиваются у дентино‑эмалевой границы, но уже на другой стороне коронки зуба.
На поперечных шлифах зубов линии Ретциуса располагаются в виде концентрических кругов, сравниваемых некоторыми авторами с годичными кольцами роста на поперечном срезе ствола дерева.
Линии Ретциуса представляют собой границы между последовательно возникающими в процессе развития зуба слоями эмали и являются участками с пониженным содержанием солей извести (зоны покоя), отражая периодичность этого процесса. В эмали 7–9 линий Ретциуса, расположенных с интервалом около 16 мкм, поэтому их формирование обусловлено ритмическим процессом с периодом около 1 нед.
Линии Ретциуса (ростовые линии эмали) наиболее отчетливо выражены в эмали постоянных зубов. Число их увеличивается при нарушениях процессов образования эмали. Так, среди этих линий иногда выделяют неонатальную линию – четко выраженную ростовую линию, соответствующую перинатальному периоду длительностью 1–1,5 нед, когда нарушается образование эмали. Эта линия определяется во всех молочных зубах и первом постоянном моляре и разделяет эмаль, образованную до и после рождения.
При световой микроскопии эмалевых призм по их ходу наблюдается также правильное чередование темных и светлых участков или полосок, которые придают исчерченный вид всей призме. Расстояние между одноименными полосками приблизительно одинаково и равно 4 мкм [Скотт Д., 1964; Дэвис В. 1986]. Эта поперечная исчерченность эмалевых призм является отражением суточного ритма отложения солей кальция в процессе развития эмалевых призм, т. е. различной интенсивности их минерализации днем и ночью.
На поперечных шлифах зуба призмы имеют овальную, гексагональную, полигональную форму или форму аркад (чаще у человека), напоминая собой рыбью чешую или замочную скважину.
Диаметр призм неодинаков: увеличивается от дентино‑эмалевой границы к поверхности эмали в 1,5–2 раза в связи с тем, что наружная поверхность эмали превышает внутреннюю, граничащую с дентином, откуда начинаются эмалевые призмы.
Эмалевые призмы состоят из плотно уложенных кристаллов гидроксиапатита и кальциевого фосфата. Каждый кристалл имеет толщину в среднем 25–40 нм, ширину 40–90 нм и длину 100–1000 нм и покрыт гидратной оболочкой с толщиной около 1 нм. Микропространства между кристаллами заполнены водой (эмалевая жидкость), служащей переносчиком ионов и молекул ряда веществ.
В центральной части каждой призмы кристаллы лежат параллельно ее длинной оси, на периферии они удаляются от этой оси, образуя с ней все больший угол. При аркадной конфигурации эмалевых призм этот угол составляет 40–65°.
Эмалевые призмы связаны между собой склеивающим межпризматическим веществом, толщиной менее 1 мкм, которое также обызвествлено, хотя и в меньшей степени, чем призмы. Аркадной формы призмы находятся в непосредственном контакте друг с другом, так что межпризматическое вещество как таковое практически отсутствует. В межпризматическом веществе кристаллы апатита имеют косую направленность по отношению к призмам, часто под прямым углом к ним.
С поверхности эмалевая призма окружена оболочкой (кора призмы), которая меньше кальцифицирована, чем остальная часть призмы, легче окрашивается и лучше сопротивляется действию кислот. Благодаря оболочке каждая призма отграничена от межпризматического вещества.
Электронно‑микроскопические исследования [Дэвис В., 1986; Тен Кате, 1994] позволили выявить неоднородность внутреннего содержимого эмалевых призм и наличие в их составе органического компонента фибриллярного строения и имеет вид тонкой белковой сеточки, равномерно пронизывающей всю призму и межпризматическое вещество. В петлях этой сети располагаются кристаллы гидроксиапатита.
Самый внутренний слой эмали толщиной 5–10 мкм у дентино‑эмалевой границы не содержит призм (начальная эмаль). В этом слое имеются мелкие кристаллы гидроксиапатита толщиной около 3–5 нм, расположенные почти перпендикулярно к поверхности эмали. Они переходят в более глубокий слой, в котором плотно расположены кристаллы размером 40–50 нм, лежащие под прямым углом к поверхности эмали. Наличие беспризменной эмали связано с отсутствием отростков Томса в период ее образования.
Аналогичным образом на завершающих этапах секреции эмали, когда у амелобластов исчезают отростки Томса, образуется наиболее наружный слой эмали (конечная эмаль), в котором также отсутствуют эмалевые призмы. Слой конечной эмали значительнее выражен в постоянных зубах, поэтому на поверхности молочных зубов преимущественно призменная структура, а поверхность постоянных зубов на большем своем протяжении гладкая.
С помощью сканирующего электронного микроскопа на поверхности эмали коронок зубов можно обнаружить и другие структуры, например отверстия диаметром 1–2 мкм в области беспризменных участков, а также микротрещины шириной 0,3–0,6 мкм, которые окружают группы в 20–30 призм, создавая в совокупности структуру в виде сот.
Эмалевые пластинки и пучки. Своеобразными структурами, присущими зрелой эмали, являются эмалевые пластинки и эмалевые пучки. Они представляют собой участки недостаточно обызвествленных эмалевых призм и межпризматического вещества, но отличаются друг от друга своей формой и положением в толще эмали.
Эмалевые пластинки – это тонкие листообразные структуры, проходящие через всю толщу эмали и содержащие белки эмали и органические вещества из полости рта. Их больше в области шейки зуба, они видны только на поперечных шлифах зуба. На последних эмалевые пластинки сходны с трещинами эмали, однако в отличие от последних заполнены органическим веществом, которое сохраняется после декальцинации.
Эмалевые пучки в виде мелких конусовидных образований, сходных внешне с пучками травы, располагаются у дентино‑эмалевой границы и в отличие от эмалевых пластинок эти пучки проникают только во внутренние отделы эмали, располагаются с интервалом 30–100 мкм. По мнению ряда авторов, как те, так и другие могут служить входными воротами для бактерий и начальными пунктами для развития кариеса.
Эмалевые веретена. Веретенообразные или булавовидные структуры, располагающиеся во внутренней трети эмали перпендикулярно к дентино‑эмалевой границе и не совпадающие по своему ходу с эмалевыми призмами. Их происхождение связывают с тем, что в период образования дентина отростки одонтобластов проходят через дентино‑эмалевое соединение, по‑видимому, для более тесной информационной связи с дифференцированными секреторными амелобластами. В процессе амелогенеза эти структуры оказываются внутри кальцинированного эмалевого вещества, сохраняясь в зрелой эмали в виде эмалевых веретен. Таким образом, структурно – это дентинные трубочки, содержащие, по В. Дэвису (1986), в зрелом зубе внеклеточную жидкость и другие органические компоненты.
Возрастные изменения эмали. С возрастом происходит постепенное стирание поверхностных слоев эмали, особенно в участках коронки, связанных с актом жевания (жевательные, резцовые поверхности зубов). Зрелая эмаль не обладает способностью к самовоспроизведению, поэтому такие потери ткани вследствие трения ничем не компенсируются. Это ведет к уменьшению вертикального размера коронки и уплощению контактных поверхностей.
Уменьшается также проницаемость эмали, что связано, с одной стороны, с увеличением в размерах кристаллов апатита и соответствующим уменьшением содержания воды в зрелой эмали, а с другой – с прогрессивным увеличением содержания фтора, а также кальция, фосфора и цинка на поверхности эмали.
Изменяется и цвет зубов. Потемнение эмали связывают не только с изменениями в структуре эмали, но и с возрастной перестройкой дентина зуба.
Дентин (dentinum) образует основную массу зуба и определяет его форму. У человека дентин в области коронки покрыт эмалью, а в области корня – цементом. Таким образом, в нормальном зубе дентин нигде не соприкасается с внешней средой и тканями, окружающими зуб.
Как и эмаль, зрелый дентин – окостеневшая ткань зуба. По своему образованию, структуре и физиологическим особенностям дентин скорее всего ближе к компактной грубоволокнистой костной ткани, но отличается от нее большей твердостью и отсутствием клеток.
Основные особенности дентина:
• он происходит из эктомезенхимы (клеток краниальной части нервного гребня);
• восстанавливается и видоизменяется в течение всей жизни индивидуума;
• это бесклеточная ткань (содержит только отростки одонтобластов), не имеющая кровеносных сосудов;
• одонтобласты участвуют и в образовании, и в минерализации органической основы дентина.
Дентин образует стенки пульпарной камеры, содержащей пульпу зуба, а также стенки корневого канала, открывающегося на верхушке корня одним или несколькими отверстиями, которые связывают пульпу с периодонтом.
Физические свойства и химический состав. Дентин имеет светло‑желтую окраску, обладает некоторой эластичностью, прочнее кости и цемента, но в 4–5 раз мягче эмали. Высокая твердость дентина связана с наличием в нем большого количества минеральных солей, содержание которых доходит до 70–80 %, в то время как остальные 20–30 % составляют органические вещества (18 %) и вода (12 %). В связи с этим в зрелом дентине выделяют органический и неорганический компоненты (матриксы).
Органический матрикс дентина состоит из коллагена – довольно крупных волоконец с аксиальной периодичностью 600–700 нм, характерных для коллагена 1‑го типа. Это генетически специфичная форма коллагена, находящаяся в наиболее окостеневших тканях (дентин, кость, цемент).
В дентине коллагеновые волокна ориентированы беспорядочно, как в плотной неоформленной соединительной ткани. Только у дентино‑эмалевой границы – в плащевом дентине, волокна характеризуются регулярностью, точной организацией и ориентацией.
Коллагеновые волокна матрикса дентина погружены в основное аморфное вещество, состоящее из таких гликозаминогликанов, как хондроитинсульфаты. Последние могут соединяться с неколлагеновыми протеинами с образованием протеогликанов – главных составляющих матрикса дентина.
Около 20 % органического матрикса дентина составляют неколлагеновые протеины, среди которых главными являются фосфопротеины, играющие определенную роль при минерализации дентина.
В органической основе дентина идентифицированы также липиды (гликолипиды и фосфолипиды), которые, вероятно, участвуют в минерализации матрикса.
Неорганический матрикс, как и в кости, цементе и эмали, состоит из фосфата кальция в форме кристаллов гидроксиапатита. Кристаллы гидроксиапатита в дентине маленькие, тонкие, иглообразные. С помощью трансмиссионного электронного микроскопа установлено, что эти кристаллы находятся как внутри, так и между коллагеновыми волокнами. В состав неорганического матрикса дентина входят также в небольшом количестве фторид кальция (фторапатит), карбонат кальция, магний и натрий.
Строение дентина. Дентин состоит из основного вещества и множества тонких дентинных канальцев, пронизывающих основное вещество.
Дентинные канальцы обеспечивают трофику дентина и представляют из себя тонкие конусообразные трубочки диаметром от 1 до 3 мкм, идущие в радиальном направлении от пульпы зуба к эмали или к цементу. Они шире во внутренних отделах дентина и постепенно суживаются кнаружи.
Количество дентинных канальцев неодинаково в различных отделах дентина. В связи с радиальным направлением канальцев по отношению к полости зуба во внутренних отделах дентина (вблизи пульпы) они лежат более тесно: на 1 мм2 дентина приходится 50–75 000 дентинных канальцев, а ближе к периферии – от 15 000 до 30 000 канальцев на 1 мм2. В коронке зуба их больше, чем в корне. В молярах на 1 мм2 поверхности дентина их приходится в 1,5 раза меньше, чем в резцах.
Дентинные канальцы в коронке S‑образно изогнуты, а в области корня зуба почти прямые и идут перпендикулярно к оси зуба.
В толще дентина канальцы ветвятся и отдают боковые ответвления, анастомозирующие между собой. Особенно ярко ветвления канальцев видны у дентино‑эмалевой и дентиноцементной границ, где каждый из канальцев делится на несколько терминальных ветвей. В некоторых случаях канальцы могут пересекать дентино‑эмалевую границу, проникая в толщу эмали и образуя эмалевые веретена.
Благодаря наличию огромного числа трубочек дентин обладает высокой проницаемостью. Это обстоятельство имеет клиническое значение, обусловливая быструю реакцию пульпы на повреждение дентина.
Окружающее канальцы основное вещество является более уплотненным (гиперминерализованным) и однородным, чем в промежутках между ними. В связи в этим выделяют перитубулярный (вокругтубулярный) и интертубулярный (межтрубочный) дентин.
Перитубулярный дентин представляет собой слой дентина, непосредственно окружающий каждую дентинную трубочку, образуя ее стенку. Толщина слоя перитубулярного дентина у пульпарного конца трубочки составляет около 40–50 нм, а у дентино‑эмалевой границы 500–700 нм. Этот дентин характеризуется повышенным (на 35–40 %) содержанием минеральных веществ по сравнению с интертубулярным дентином. Содержание органических веществ в перитубулярном дентине минимально – при декальцинации он почти полностью исчезает.
Интертубулярный дентин в процессе развития зуба образуется первым как в плащевом, так и в околопульпарном дентине. Он состоит в основном из обызвествленных коллагеновых фибрилл диаметром 100–200 нм, при этом кристаллы гидроксиапатита расположены вдоль оси фибрилл.
Содержимое дентинных трубочек разнообразно: отростки одонтобластов (отростки Томса), безмиелиновые нервные волокна, тканевая дентинная жидкость, необызвествленные коллагеновые фибриллы (интратубулярные фибриллы), кристаллы гидроксиапатита.
Изнутри стенка дентинной трубочки выстлана тонкой пленкой органического вещества – пограничной пластинкой (мембрана Неймана). Она проходит по всей длине дентинного канальца и содержит большое количество гликозаминогликанов.
Отростки одонтобластов, являясь непосредственным продолжением апикальных отделов их клеточных тел, как правило, тянутся по всей длине дентинных трубочек, заканчиваясь у дентино‑эмалевой границы. Они содержат мало органелл, но значительное количество элементов цитоскелета, а также пузырьков, лизосом и полиморфных вакуолей.
Боковые ветви отростков многочисленны в предентине и внутренних отделах дентина (в пределах 100–200 мкм от границы с пульпой), они редки в средних его отделах и вновь часто встречаются на периферии. Ветви образуют контакты с ответвлениями соседних одонтобластов, что играет существенную роль в передаче питательных веществ и ионов, а также способствует распространению микроорганизмов и кислот при кариесе.
Нервные волокна, направляющиеся в дентин из периферических отделов пульпы, проникают в него обычно на глубину нескольких микрометров, отдельные волокна – на 100–200 мкм.
Часть волокон уже в предентине делится на многочисленные ветви. Другие проходят внутри дентинных канальцев вдоль отростка одонтобласта или имеют спиральный ход, оплетая его и изредка формируя ответвления, идущие под прямым углом к трубочкам.
Основное вещество дентина, расположенное между дентинными канальцами, имеет фибриллярную структуру и состоит из коллагеновых волокон и однородного цементирующего их вещества. Расположение волокон и их структура различны в разных отделах дентина. В связи с этим различают два слоя дентина: наружный, или плащевой (мантийный), дентин и внутренний, или околопульпарный, дентин.
В плащевом дентине преобладают волокна, идущие в радиальном направлении (волокна Корфа). В общем волокна Корфа расположены параллельно ходу канальцев. Это расположение сохраняется только в области верхушки коронки зуба, а на боковых поверхностях коронки и в области корня они приобретают все более косое направление.
Плащевой дентин нерезко переходит в околопульпарный, причем к радиальным волокнам примешивается все большее количество тангенциальных. Матрикс плащевого дентина менее минерализован, чем матрикс околопульпарного, и содержит относительно меньше коллагеновых волокон.
В околопульпарном дентине волокна располагаются тангенциально (волокна Эбнера), т. е. параллельно поверхности пульпы и приблизительно под прямым углом к дентинным канальцам. Расположение этих волокон в околопульпарном дентине совпадает с расположением дентинных пластинок, т. е. слоев дентина, отлагающихся изнутри, со стороны пульпы в процессе развития зуба. Выражением ритмичности роста и слоистого отложения дентина являются линии роста – контурные линии Оуэна и ростовые линии Эбнера.
Линии Оуэна идут обычно под прямым углом к дентинным канальцам и соответствуют периодам покоя одонтобластов, что сопровождается менее полным обызвествлением вещества дентина и образованием в этих местах очень мелких интерглобулярных пространств.
В молочных и первых постоянных коренных зубах нередко видна контурная (неонатальная) линия, отделяющая слой дентина, образовавшийся в период эмбриональной жизни, от дентина, который возник после рождения. По мнению Б. Орбана (1974), эта линия соответствует неполностью обызвествленному дентину, возникшему в первые 2 нед после рождения в связи с расстройством обмена в период приспособления новорожденного к резким изменениям среды и питания.
Более тонкие линии Эбнера, направленные почти перпендикулярно к дентинным трубочкам, располагаются ближе друг к другу, чем линии Оуэна, с периодичностью в коронке около 18–20 мкм. Между линиями Эбнера имеются линии, расстояние между которыми обычно постоянное, около 4–5 мкм, в зависимости от положения в верхушке или корне зуба. Предполагают, что последние отражают дневную ритмичность в образовании дентина, а линии Эбнера соответствуют 5‑суточному циклу отложения органического матрикса дентина, т. е. периодам меньшей минерализации основного вещества дентина.
Вторичный и третичный дентин. Дентин, отлагающийся в зубах взрослого человека в течение его жизни, называется вторичным (регулярным, или физиологическим вторичным) дентином. Помимо более медленного темпа образования, он отличается от первичного дентина (возникшего в процессе эмбрионального развития зуба) менее правильной структурой. Это выражается в изменении хода и числа дентинных канальцев и коллагеновых волокон, более низкой степенью минерализации.
Трубочки вторичного дентина располагаются реже, они более узкие; пересекая границу первичного и вторичного дентина (демаркационную линию) могут S‑образно изгибаться.
Наиболее активно отложение вторичного дентина происходит в боковых стенках и в крыше пульпарной камеры, а в многокорневых зубах – в ее дне. В связи с этим с возрастом форма пульпарной камеры изменяется (сглаживаются, в частности, рога пульпы), а объем ее уменьшается.
Интенсивность отложения вторичного дентина у мужчин выше, чем у женщин; с возрастом она снижается. Толщину слоя вторичного дентина можно использовать в качестве одного из показателей для оценки возраста индивидуума.
Продукция вторичного дентина резко усиливается при разрушении или стирании эмали и обнажении дентина (кариес, повышенная стираемость зубов, воздействие химических веществ и т. д.). В участках пульпы, соответствующих области повреждения зуба, отмечается отложение более или менее значительных масс заместительного дентина, которые могут вдаваться в полость зуба и изменять ее конфигурацию. Такой дентин называют третичным, или иррегулярным.
В отличие от вторичного дентина, который выстилает пульпу, образование третичного дентина происходит более или менее локально, т. е. только в местах наиболее сильного воздействия неблагоприятного фактора. В связи с этим основной функцией третичного дентина является защита пульпы зуба от распространения внутрь вредных веществ (бактерии, токсины и т. п.). Он может образовываться в любом участке стенки пульпарной камеры, но наиболее часто – в области рогов пульпы.
Дентикли. В пульпе зуба иногда наблюдаются тела округлой или неправильной формы, состоящие из дентина или дентино‑подобной ткани. Они получили название дентиклей, или камней пульпы. По своему положению в пульпе они делятся на свободные, т. е. лежащие непосредственно в пульпе; пристеночные, сохраняющие связь со стенкой зуба; интерстициальные, которые возникают при обрастании свободного или пристеночного дентикля новыми слоями вторичного дентина. В результате дентикль оказывается замурованным в стенке зуба (этот процесс чаще наблюдается в корне зуба, ближе к его верхушке).
В зависимости от структуры принято различать высокоорганизованные (канализированные) и низкоорганизованные (лишенные канальцев) дентикли. Источником их образования являются одонтобласты. Дентикли имеют строение дентина или дентиноподобной ткани, поэтому они называются истинными дентиклями в отличие от ложных дентиклей, представляющих собой очаги ограниченного обызвествления в ткани пульпы.
Размеры дентиклей весьма непостоянны – от еле заметных зерен до 2–3 мм в диаметре. В результате своего роста дентикли могут сливаться друг с другом, заполняя собой всю пульпарную камеру или каналы корней, препятствуя их обработке. Сдавливая нервные стволики пульпы, дентикли могут вызывать боли пульпитного характера.
Возрастные изменения дентина. В отличие от эмали, дентин – живая ткань, продуцируемая непрерывно на протяжении жизни человека. Образование вторичного, а затем третичного дентина с возрастом неуклонно ведет к уменьшению размеров, объема и формы пульпарной камеры – процессу, называемому клиницистами рецессией пульпы.
В зубах пожилых людей наблюдаются участки дентина, в которых соли извести откладываются не только в основном веществе, но и в дентинных канальцах и отростках дегенерирующих одонтобластов. В результате происходит облитерация («физиологический» склероз), т. е. полное закрытие просвета некоторых групп дентинных канальцев, при этом показатели преломления канальцев и основного вещества выравниваются и поэтому такие участки кажутся прозрачными. Их называют склеротическим (прозрачным) дентином. Образование его происходит также при кариесе или повышенной стираемости зубов («патологический» склероз), что можно рассматривать как защитную реакцию зуба на действие вредного агента, предохраняющую пульпу от раздражения и проникновения в нее инфекции.
Вследствие того что склерозирование дентина снижает его проницаемость, оно может продлить период жизнеспособности пульпы. Склерозирование трубочек снижает чувствительность зуба.
С возрастом, особенно при повышенной стираемости зубов, нередко наблюдаются гибель части одонтобластов и их отростков и закупорка внутренних концов соответствующих дентинных канальцев третичным дентином. Содержимое таких канальцев распадается, а полости канальцев заполняются воздухом или другими газообразными веществами. Вследствие этого на шлифах зуба группы таких канальцев кажутся черными в проходящем свете. Фиш (1932) назвал группы таких канальцев мертвыми путями. Чувствительность дентина в этих участках снижена. В большинстве случаев «мертвые пути» со стороны пульпы, как отмечалось, закрываются путем образования репаративного третичного дентина.
Пульпа зуба (pulpa dentis) заполняет полость зуба в области как коронки, так и его корня, повторяя внешний рельеф зуба. В коронке она образует выступы, соответствующие бугоркам жевательной поверхности, – рога пульпы. Пульпа почти полностью окружена твердой тканью – дентином, благодаря чему является довольно уникальной тканью, похожей на костный мозг.
Основные функции пульпы:
• трофическая – питание дентина (за счет находящихся в ней сосудов);
• сенсорная (вследствие наличия в ней большого количества нервных окончаний);
• защитная (путем клеточных и гуморальных реакций, воспаления);
• пластическая и репаративная (путем выработки первичного, вторичного и третичного дентина).
В целом пульпа представляет собой обильно кровоснабжаемую и иннервируемую специализированную рыхлую неоформленную соединительную ткань, содержащую клетки, волокна и аморфное основное вещество. В некоторых отделах зуба эти компоненты организованы в четкие демаркационные зоны.
Пульпа коронки зуба содержит на единицу площади больше клеток и межклеточного вещества, чем коллагеновых волокон. Клетки разнообразные, хорошо дифференцированные. Одонтобласты в этой части пульпы имеют призматическую форму и располагаются в несколько рядов.
Пульпа, заполняющая корневые каналы зуба (пульпа корня), построена по типу довольно плотной соединительной ткани с преобладанием пучков коллагеновых волокон над клеточными элементами. Она слабее васкуляризована и иннервирована, чем коронковая, ее клеточный состав менее разнообразен; одонтобласты кубической или уплощенной формы, располагаются обычно в 1–2 ряда. Эти различия в структуре зависят, по‑видимому, от особенностей питания твердых тканей зуба в области коронки и корня.
В коронковой пульпе различают 4 зоны по составу и свойствам ткани [Дэвис В., 1986]: 1) одонтобластическую; 2) светлую зону Вейля; 3) субодонтобластическую; 4) центральную зону.
В самой наружной одонтобластической зоне компактно располагаются в один или несколько рядов вытянутые клетки с базофильной цитоплазмой – одонтобласты. Отростки этих клеток (отростки Томса) проникают в дентинные канальцы и пронизывают всю толщу дентина. Одонтобласты тесно связаны друг с другом межклеточными соединениями, между которыми проходят нервные волокна, направляющиеся вместе с отростками одонтобластов в дентинные трубочки.
Одонтобласты и их отростки играют важную роль в питании зуба и доставке минеральных солей к эмали и дентину. Они сохраняются в пульпе зуба взрослого человека в течение всей его жизни, при этом они выполняют и свою дентинообразующую функцию, хотя не так интенсивно, как это имело место в период развития зуба.
За слоем одонтобластов располагается бедный клетками светлый слой Вейля, состоящий главным образом из коллагеновых и ретикулярных волокон, а также отростков клеток субодонтобластической зоны. Он хорошо выражен в коронковой пульпе и отсутствует в корневой. Слой Вейля образуется довольно поздно, часто уже после прорезывания зубов. Современные исследования показали, что в этой зоне располагаются обширные сети нервных волокон и кровеносных капилляров. От нервного сплетения (сплетение Рашкова) берут начало ветви, образующие древовидные разветвления на одонтобластах, а также проходящие между ними в дентинные трубочки, где они контактируют с отростками Томса.
Третий слой пульпы – субодонтобластическая зона состоит из большого количества звездчатых клеток (преодонтобластов), которые относят к малодифференцированным клеткам. От тел этих клеток отходят многочисленные тонкие и длинные отростки, они многократно ветвятся и анастомозируют между собой. По мнению ряда авторов (Г. Ясвоин, Л. Фалин и др.), эти клетки способны к дифференцировке и превращению в одонтобласты, а также в фибробласты. Во взрослом зубе при гибели части одонтобластов возможна их замена за счет дифференцировки звездчатых клеток субодонтобластического слоя. Кроме того, в этой зоне располагаются также фибробласты, лимфоциты, макрофаги, малодифференцированные клетки, а также капилляры, миелиновые и безмиелиновые нервные волокна.
Центральная зона пульпы представляет собой рыхлую волокнистую соединительную ткань, богатую клетками, сосудами и нервами. В пульпе зрелого зуба постоянно выявляются четыре основные клеточные группы: 1) одонтобласты; 2) фибробласты; 3) малодифференцированные эктомезенхимальные клетки; 4) макрофаги.
Кроме того, в меньшем числе постоянно присутствуют дендритные клетки (фиксированные макрофаги), лимфоциты, плазматические, тучные клетки, гранулоциты крови.
Одонтобласты – самая большая клеточная популяция в пульпе. Это специфические клетки пульпы, которые образуют дентин и обеспечивают его трофику. Одонтобласты располагаются на периферии пульпы очень плотно в коронковой пульпе (40–50 000 на 1 мм2 поверхности дентинопульпарной границы) и реже (15–20 000 на 1 мм2) в корневой пульпе. Клетки призматические или грушевидные в пульпе коронки, ближе к корню они веретеновидные, а в пульпе корня кубические или уплощенные. Форма клетки изменяется не только в зависимости от ее расположения, но и в связи с функцией: чем выше активность, тем большую высоту имеет одонтобласт.
В призматических клетках четко выявляется овальное ядро с 1–2 ядрышками, расположенное в базальной части; в кубических одонтобластах сферическое ядро лежит обычно центрально. В ядрах одонтобластов преобладает эухроматин, только у ядерной оболочки отмечаются небольшие скопления гетерохроматина. В цитоплазме располагаются цистерны гранулярной ЭПС, большое число рибосом, лизосом, митохондрий, гранулы гликогена, мелкие липидные капли, а в апикальной части – секреторные гранулы, содержащие преколлаген и протеогликаны предентина.
Апикальная часть тела одонтобласта суживается, продолжаясь в длинный ветвящийся отросток Томса, который направляется в дентинную трубочку. Отросток характеризуется хорошо развитыми элементами цитоскелета, ориентированными продольно; многочисленными гладкими и окаймленными пузырьками, редкими митохондриями и короткими цистернами агранулярной ЭПС.
Соседние одонтобласты прочно связаны межклеточными соединениями (десмосомами, щелевыми и плотными контактами), благодаря которым слой одонтобластов способен выполнять барьерную функцию, регулируя перемещение молекул и ионов между пульпой и предентином.
Одонтобласты относят к окончательно дифференцированным клеткам, поэтому продолжительность их жизни может достигать длительности существования зуба (Быков В. Л., 1996).
Фибробласты находятся в большом количестве в центральной зоне пульпы, особенно у молодых людей. Они берут начало от недифференцированных эктомезенхимальных клеток, также расположенных в пульпе. Фибробласты могут существовать в одном из двух функциональных состояний: активном (фибробласты) или пассивном (фиброциты). Функциональное состояние клеток зависит от возраста, клинических характеристик пульпы и т. п.
Структурно фибробласты пульпы – это веретенообразные отростчатые клетки со светлым ядром с мелкодисперсным хроматином и крупным ядрышком, слабобазофильной цитоплазмой, хорошо развитыми гранулярной ЭПС и комплексом Гольджи, большим количеством везикул, микротрубочек, микрофиламентов и пиноцитозных пузырьков. Главный продукт деятельности фибробластов пульпы – коллагены I и III типов. Они же синтезируют и выделяют компоненты основного аморфного вещества пульпы – гликозаминогликаны, протеогликаны, гликопротеины. При воспалительных процессах (пульпит) фибробласты принимают участие в образовании фиброзной капсулы, ограничивающей очаг воспаления. Помимо синтетической активности, они способны поглощать и переваривать компоненты межклеточного вещества.
Альтернативно фиброцит – это неактивная, отдыхающая клетка, со слаборазвитыми органеллами синтеза белка. В принципе это два состояния одной и той же клетки, которые проявляются в зависимости от состояния окружающей соединительной ткани. Высокая синтетическая активность свойственна фибробластам зубов молодых людей; с возрастом увеличивается доля клеток с низкими пластическими характеристиками. Описана III стадия – так называемый фиброкласт, или клетка, резорбирующая коллаген. Наличие фиброкластов впервые замечено в периодонтальной связке, хотя их присутствие в дентальной пульпе пока не установлено, некоторые исследователи считают это вопросом времени.
Другим основным типом клеток коронковой пульпы человека являются недифференцированные эктомезенхимальные клетки. Это клетки треугольной формы с большим ядром, содержащим значительное количество гетерохроматина. В цитоплазме – большое количество рибосом и полирибосом, липидных капель и хорошо развитая гранулярная ЭПС. Отмечается также большое количество микрофиламентов и микротрубочек, что указывает на достаточную подвижность этих клеток. Клетки обладают плотным гликокаликсом, содержащим мембранные рецепторы, антигенные зоны и т. п., которые функционируют в механизмах узнавания клеток и их скоплений. Во многих случаях эти клетки соединены между собой посредством филоподий и микроотростков в сложную сеть.
Большая популяция недифференцированных эктомезенхимальных клеток в пульпе зуба указывает на ее высокую регенеративную способность.
При воздействии определенных возбудителей эти клетки могут претерпевать окончательную дифференцировку либо в фибробласты, либо в одонтобласты пульпы. Последние замещают старые или погибшие клетки, а также формируют одонтобласты, необходимые для образования вторичного и третичного дентина. С возрастом отмечается уменьшение количества недифференцированых клеток пульпы, что ведет к снижению способности пульпы к регенерации при старении.
В отличие от трех описанных типов клеток центральной зоны пульпы, имеющих общее происхождение от клеток головной части нервного гребешка, макрофаги берут начало от циркулирующих специфических кровяных клеток – моноцитов. Их количество в пульпе, морфология и метаболизм зависят от функционального состояния окружающей соединительной ткани. Особенно многочисленны макрофаги в пульпе зубов молодых людей. Это полиморфные клетки с эксцентрично расположенным ядром. В цитоплазме находятся хорошо развитые гранулярная ЭПС и комплекс Гольджи, а также большое количество лизосом и вакуолей. Хорошо развитый цитоскелет из микронитей и микротрубочек обеспечивает высокую подвижность этих клеток. Поверхность клетки покрыта множеством складок и выростов, указывающих на ее выраженную фагоцитирующую активность.
Наличие в пульпе зуба макрофагов показывает, что она, помимо своей трофической и дентинообразующей функции, выполняет еще и важную защитную или барьерную роль.
Дендритные клетки являются также постоянным компонентом пульпы, составляя 6–8 % от общей клеточной популяции пульпы. Это клетки вариабельной формы с многочисленными ветвящимися отростками, четко выраженным ядром и хорошо развитым лизосомальным аппаратом. Они являются антиген‑представляющими клетками, т. е. их функция заключается в поглощении различных антигенов, их процессинге и представлении лимфоцитам. По способности индуцировать пролиферацию Т‑лимфоцитов дендритные клетки намного превосходят макрофаги, находясь с последними в пульпе в соотношении 4:1. Число этих клеток резко возрастает при антигенной стимуляции.
Больше всего дендритных клеток в рогах коронковой пульпы. Они располагаются вдоль сосудов, около одонтобластов и в субодонтобластическом слое. Содержание их в пульпе невелико после рождения, но увеличивается по мере созревания пульпы.
Лимфоциты, преимущественно, малые лимфоциты (около 90 %), присутствуют в пульпе в небольшом количестве в периферических ее участках. Это чаще всего различные субпопуляции Т‑клеток. В‑лимфоциты в нормальной пульпе почти не обнаруживаются. Наблюдаются единичные плазматические клетки, которые являются конечными стадиями дифференцировки В‑лимфоцитов. Эти клетки активно синтезируют иммуноглобулины, обеспечивая реакции гуморального иммунитета.
Следует отметить, что содержание лимфоцитов в пульпе резко возрастает при воспалении. В этот же период в пульпе выявляются тучные клетки, содержащие биологически активные вещества (гепарин, гистамин, эозинофильный хемотаксический фактор, лейкотриены), выделение которых вызывает ряд эффектов, в том числе расширение сосудов и увеличение проницаемости их стенки. По мнению некоторых авторов, тучные клетки присутствуют в пульпе постоянно у детей.
Кроме описанных видов клеток, в пульпе отмечаются и другие клетки. Большинство этих клеток находится в соединении с сосудистой стенкой: эндотелиальные клетки, перициты, гладко‑мышечные клетки, а также редкие эозинофильные гранулоциты.
Центральная пульпа содержит также коллагеновые волокна. Коллаген составляет 25–30 % от сухой массы пульпы зуба, при этом его содержание с возрастом увеличивается. Современные биохимические исследования показали, что это коллаген I и III типов.
Собственно коллагеновые волокна (коллаген I типа) создают систему поддерживающих элементов для клеток, кровеносных сосудов и нервов. В центральных отделах коронковой пульпы они располагаются рыхло, в периферических – образуют более плотные скопления. В корневом канале волокна ориентированы по его длине, образуя плотные пучки. Часть волокон проникает между одонтобластами, смешиваясь с волокнами предентина.
По всей пульпе в виде сети располагаются ретикулярные волокна (образованные коллагеном III типа). В процессе образования дентина эти многочисленные волокна лежат первоначально между одонтобластами и называются волокнами Корфа.
В пульпе выявлены также преколлагеновые, окситалановые и эластические волокна. Последние являются, строго говоря, компонентами стенки кровеносных сосудов. Окситалановые волокна многочисленны в периферической части пульпы, не имеют строгой ориентации и связаны обычно с кровеносными сосудами. Их относят к преэластическим волокнам.
Как указано, пульпа, помимо клеток и волокон, содержит тонкое аморфное вещество, которое является компонентом внеклеточного матрикса пульпы. Оно состоит из воды, гликозаминогликанов (хондроитинсульфаты и гиалуроновая кислота), гликопротеинов, протеогликанов.
Обладая высокими способностями к диффузии, аморфное межклеточное вещество способствует распространению питательных веществ между клетками, сосудами и нервами.
Кровоснабжение пульпы. Васкуляризация и ангиоархитектоника сосудов в пульпе имеют свои особенности. Все кровеносные и лимфатические сосуды, а также нервы входят (и выходят) в пульпу в основном через отверстие в верхушке корня зуба, образуя в корневом канале сосудисто‑нервный пучок. В некоторых случаях дополнительные сосуды и другие структуры входят в пульпу через добавочные каналы боковых стенок корня зуба. Эти каналы имеют клиническое значение, поскольку могут способствовать распространению инфекции из периодонта в пульпу и обратно.
После входа в корневую пульпу сосуды – артериолы диаметром 50–150 мкм имеют почти прямую (вертикальную) направленность в коронковую пульпу, занимая ее центральную зону. От центрального сосуда отходят более мелкие прекапиллярные артериолы диаметром 10–12 мкм, которые образуют затем в слое Вейля обширное капиллярное сплетение, питающее пульпу и одонтобласты. Капилляры пульпы диаметром 8–10 мкм делятся на две группы: фенестрированные (около трети), располагающиеся в основном вблизи одонтобластов, и соматические (2/3 всех капилляров). Последние характеризуются наличием множества пиноцитозных пузырьков в цитоплазме эндотелия сосудов. Как те, так и другие обеспечивают питанием все структурные элементы пульпы зуба.
Объем капиллярного русла в пульпе может значительно варьировать, чему способствует наличие артериоловенулярных анастомозов. В состоянии покоя большая часть анастомозов и капилляров не функционирует, однако их деятельность резко усиливается при раздражении пульпы, приводя к развитию гиперемической реакции. С этим механизмом также связывают периодичность болей при пульпите.
От субодонтобластической капиллярной сети кровь поступает в венулы, которые постепенно сливаются в две‑три более крупные венулы мышечного типа (содержат в стенке гладкие миоциты). Как правило, венулы располагаются в пульпе центрально, тогда как артериолы занимают более периферическое положение. Уникальной особенностью кровеносной системы пульпы является то, что диаметр выносящих венул меньше диаметра входящих в пульпу артериол. В связи с этим даже в нормальной пульпе всегда наблюдаются явления гиперемии и стаза крови, что позволило Г. Фишеру (1995) назвать пульпу застойным органом. Давление в пульпарной камере составляет 20–30 мм рт. ст., что значительно выше внутритканевого давления в других органах. Указанные структурные особенности сосудистой системы, по‑видимому, обеспечивают медленный кровоток через пульпу, что имеет определенное физиологическое значение для питания тканей зуба.
Вопрос о лимфатических сосудах пульпы во многом остается открытым. Одни исследователи считают, что в пульпе имеются лимфатические капилляры и сосуды, другие отмечают наличие тканевых пространств и щелей, выстланных эндотелием, которые сообщаются с венозными сосудами. Отток лимфы от пульпы происходит в поднижнечелюстные и подподбородочные лимфатические узлы.
Иннервация пульпы. Достаточно крупные нервные миелиновые и безмиелиновые волокна тройничного нерва входят в пульпу через отверстие в верхушке корня вместе с кровеносными сосудами, формируя в корневом канале сосудисто‑нервный пучок, обеспечивающий питание и иннервацию зуба. Безмиелиновые волокна составляют до 60–80 % от общего числа волокон. В корневой пульпе лишь около 10 % волокон образуют терминальные ветвления, большая их часть достигает коронки, где они веерообразно расходятся к периферии пульпы. В пульпе коронки они обильно ветвятся, формируя в слое Вейля субодонтобластическое нервное сплетение (сплетение Рашкова), содержащее как толстые миелиновые, так и тонкие безмиелиновые волокна. От сплетения отходят ветви к одонтобластам и в дентинные трубочки между ними. Одни из них формируют окончания на телах одонтобластов, другие на одонтобластических отростках Томса, проникая, однако, только на 1/3 толщины дентина зрелого зуба.
Большинство нервных окончаний в области тел одонтобластов относят к рецепторам. Раздражение этих рецепторов независимо от природы действующего фактора (давление, холод, тепло, химические вещества) вызывают болевые ощущения. В последнее время описаны и эффекторные окончания в пульпе зрелого зуба.
Среди миелиновых волокон пульпы выделяют более многочисленные α‑волокна диаметром 1–6 мкм, которые опосредуют болевую чувствительность, в частности ощущение острой локализованной боли, и β‑волокна диаметром 6–12 мкм, обеспечивающие, предположительно, проведение тактильных сигналов.
Безмиелиновые волокна диаметром 0,5–1,5 мкм также подразделяются на несколько групп. Часть волокон является преимущественно вазомоторными, регулируя тонус артериол и кровоток в пульпе, другие содержат нейропептиды (энкефалины, холецистокинин, соматостатин, вазоинтестинальный полипептид и др.) и, будучи связаны с кровеносными сосудами и одонтобластами, принимают участие в регуляции кровотока в пульпе, обеспечивают болевую чувствительность, а также влияют на развитие воспаления.
Возрастные изменения. С возрастом, в связи с непрерывным отложением вторичного и периодическим отложением третичного дентина, объем пульпарной камеры уменьшается, что ведет к изменению ее формы (сглаживаются рога пульпы), а также структурным и объемным соотношениям всех элементов пульпы.
В пульпе снижается (до 50 % от исходного) содержание клеточных элементов (фибробласты и недифференцированные эктомезенхимальные клетки), уменьшается количество преколлагеновых и почти в 3 раза увеличивается число зрелых коллагеновых волокон, уменьшается содержание жидкости в межклеточном аморфном основном веществе пульпы.
За счет возрастного сужения апикальной части канала корня зуба уменьшается васкуляризация пульпы в связи с редукцией микроциркуляторного русла, что, несомненно, влияет на жизнеспособность одонтобластов. Сокращается число рядов одонтобластов, в них становится меньше органелл, участвующих в синтетических процессах, и секреторных гранул, усиливаются аутофагические процессы.
С возрастом снижается синтетическая активность клеток фибробластического ряда, уменьшается число безмиелиновых волокон, происходят демиелинизация и гибель миелиновых волокон, а также нервных окончаний в пульпе, что влияет на чувствительность дентина и сказывается на регуляции кровоснабжения пульпы. Кроме того, увеличивается частота формирования в пульпе диффузных отложений солей извести (петрификаты) вдоль крупных кровеносных сосудов и нервов или независимо от них, что ведет иногда к образованию камней пульпы.
Цемент (cementum) покрывает дентин корня на всем протяжении – от шейки зуба и до верхушки корня, где он достигает наибольшей толщины. Это специализированная кальцифицированная соединительная ткань, напоминающая по строению грубоволокнистую кость. Цемент в отличие от кости не имеет сосудов, нервных окончаний, не испытывает в норме резорбции и перестройки структуры с возрастом.
Основной функцией цемента является связывание различных коллагеновых волокон из периодонтальной связки с веществом зуба. В этом состоит важная роль цемента – укрепление зуба в костной альвеоле челюсти.
Цемент растет путем присоединения новых слоев к старым в течение всего периода существования зуба, обеспечивая, таким образом, постоянно существующий участок для присоединения новых волокон периодонтальной связки. Это очень важно, поскольку коллагеновые волокна периодонта подвергаются в зависимости от состояния зуба почти непрерывному процессу перестройки. Кроме того, непрерывный прирост толщины цемента, особенно в апикальной области, сохраняет нормальную длину зуба, компенсируя, таким образом, уменьшение с возрастом толщины эмали.
Цемент защищает дентин корня от повреждающих воздействий, выполняет репаративные функции при образовании резорбционных лакун или переломе корня.
Цемент – это минерализованная соединительная ткань, и, следовательно, имеет как органическую, так и неорганическую фазу.
Неорганическая фаза составляет 50–60 % (по массе) зрелого цемента. Она состоит из игольчатых кристаллов гидроксиапатита кальция. Органическая фаза представлена в основном коллагеном I типа, составляющим от 40 до 50 % сухой массы. Кроме того, в нее входят различные неколлагеновые протеогликаны и гликопротеины, фосфопротеины, фосфаты.
Коллаген цемента происходит из двух различных источников: цементобластов и фибробластов периодонта.
Цементобласты синтезируют и выделяют так называемые внутренние коллагеновые волокна, тогда как фибробласты продуцируют внешние коллагеновые волокна.
Внутренние, меньшего диаметра, коллагеновые волокна составляют собственные волокна цемента, идущие обычно параллельно его поверхности. Внешние, большего диаметра, коллагеновые волокна возникают в периодонте и проникают в цемент в качестве волокон Шарпи, обычно под прямым углом к его поверхности. Места внедрения в цемент волокон периодонта имеют вид кратеров, расположенных в центре куполообразных участков цемента диаметром 6–10 мкм, приподнятых над его поверхностью, которые в совокупности занимают до 30–40 % от поверхности корня зуба, покрытой цементом. С противоположного конца они продолжаются в пучки коллагеновых волокон периодонта, которые в свою очередь переходят в шарпеевские волокна альвеолярной кости. Это обусловливает прочное укрепление корней зубов в альвеолах.
Оба типа коллагеновых волокон состоят из коллагена I типа, хотя есть данные, что внешние коллагеновые волокна могут иметь коллаген III типа, связанный с ними. В целом цемент, расположенный ближе к соединению цемент – эмаль, имеет больше коллагеновых волокон, чем тот, что расположен у верхушки корня зуба.
Большинство внешних волокон, входящих в цемент, минерализуются и становятся неотличимыми от внутренних волокон в межклеточном веществе цемента. В области апикального цемента могут располагаться и необызвествленные волокна.
Цемент, покрывающий в виде тонкого слоя боковые поверхности корня, обычно не содержит клеток. Это так называемый бесклеточный, или первичный, цемент, толщиной 30–50 мкм или менее. Эта ткань характеризуется наличием множества возрастных линий, ориентированных параллельно поверхности корня, располагающихся обычно близко друг к другу. Здесь же присутствует множество внутренних и внешних коллагеновых волокон, полностью минерализованных и практически неотличимых друг от друга. Бесклеточный цемент обладает малой скоростью роста. Он является единственным слоем цемента, покрывающим шейку зуба, а в некоторых зубах (в передних резцах нижней челюсти) он почти целиком покрывает корень. Граница бесклеточного цемента с дентином выражена нечетко.
Цемент, расположенный у верхушки корня, а в многокорневых зубах – в межкорневых отделах, содержит большое количество отростчатых клеток – цементоцитов (цементные тельца), которые находятся обычно в костных полостях, и цементобластов, лежащих на поверхности цемента. Этот цемент называется клеточным, или вторичным (толщина его 200–600 мкм). Он располагается поверх бесклеточного цемента, однако иногда при отсутствии последнего непосредственно прилежит к дентину. Граница клеточного цемента с дентином, как правило, выражена отчетливо.
Обычно цемент не содержит гаверсовых каналов и кровеносных сосудов, питание его осуществляется диффузно со стороны перицемента. Важную роль в этом играет система канальцев, отходящих от цементоцитов и содержащих отростки этих клеток.
В отличие от внеклеточного клеточный цемент растет очень быстро, при этом цементобласты превращаются в цементоциты, полностью окруженные отвердевшим матриксом. Цементоциты по строению сходны с остеоцитами. Это уплощенные клетки с крупным ядром и умеренно развитыми органеллами. От их тел отходят многочисленные (до 30) ветвящиеся отростки, которые располагаются в канальцах, ориентированных преимущественно в сторону периодонтальной связки, т. е. к источнику питания. Когда клеточный цемент становится особенно толстым, самые глубокие цементоциты часто погибают, оставляя пустые или заполненные клеточным детритом лакуны. Напротив, ближе к поверхности цемента цементоциты в большей степени сохраняют функциональную активность и сходство с цементобластами.
Цементобласты, напротив, – активные клетки с хорошо развитым синтетическим аппаратом, обеспечивающим ритмическое отложение новых слоев цемента. Располагаясь обычно в периферических отделах периодонта вокруг корня зуба, цементобласты при формировании бесклеточного цемента отодвигаются кнаружи от отложенного слоя, а при образовании клеточного цемента замуровываются в нем, превращаясь в цементоциты.
Матрикс клеточного цемента состоит в основном из минерализованных внутренних волокон. Здесь значительно меньше внешних волокон, эти волокна часто только частично обызвествлены в периферических отделах. Центральные части этих волокон остаются необызвествленными.
Соединение цемент – эмаль. В трети случаев внеклеточный цемент и эмаль встречаются точно на шеечной линии и приблизительно в 2/3 – цемент частично покрывает эмаль. В 10 % случаев между цементом и эмалью имеется зазор, обнажая, таким образом, дентин корня на шеечной границе. Исследования с помощью сканирующего электронного микроскопа показали, что и в этом случае дентин все же покрыт очень тонким слоем цемента.
Возрастные изменения. С возрастом происходит постепенное утолщение цемента, хотя скорость отложения снижается. Отложение клеточного цемента, который гораздо толще внеклеточного, может привести к постепенному сужению каналов корней и нарушению кровоснабжения и иннервации пульпы зуба. С другой стороны, непрерывное прирастание апикального цемента способствует сохранению общей длины зуба, компенсируя стирание коронки в результате изнашивания эмали. Эта компенсаторная реакция, направленная на поддержание размеров клинической коронки, называется пассивным прорезыванием зуба. Таким образом, толщина цемента позволяет оценить возраст зуба, хотя на нее могут влиять и другие факторы (например, болезни периодонта), и может быть использована в судебно‑медицинских, антропологических и археологических исследованиях.
Периодонт (periodontium). Термин «периодонт» (перицемент) употребляется чаще, чем термин «периодонтальная связка». Это важная часть поддерживающего аппарата зуба, расположенная между цементом и стенкой альвеолы. Функции периодонта разнообразны: опорно‑удерживающая, распределяющая давление, трофическая, барьерная, пластическая, сенсорная, гемостатическая и др.
По строению периодонт – это соединительная ткань, богатая коллагеновыми волокнами и клетками, объединенными в единый связывающий комплекс внеклеточным аморфным веществом. Клетки периодонтальной связки вследствие своих синтетических функций отвечают за сохранение периодонта, цемента и альвеолярной кости.
Коллагеновые волокна, состоящие из коллагена I и II типов, одним концом вплетаются в цемент, другим – в стенки костной альвеолы, образуя пучки со специфической ориентацией, поддерживающие зуб в альвеоле.
Волокна натянуты в узкой щели шириной 0,2–0,3 мм, ограниченной корнем зуба и стенкой костной альвеолы, которая называется периодонтальным пространством. Ширина этого пространства не остается постоянной: она уменьшается при бездействии зуба и увеличивается при интенсивных окклюзионных нагрузках. Две трети объема периодонтального пространства занимают коллагеновые пучки, остальную часть – рыхлая волокнистая соединительная (интерстициальная) ткань.
Основное внеклеточное вещество занимает до 65 % от объема межклеточного пространства и содержит гликозаминогликаны (преимущественно дерматансульфат) и гликопротеины. По строению оно представляет собой очень вязкий гель, содержащий до 70 % воды, благодаря чему оно играет существенную роль в амортизации нагрузок, воздействующих на зуб во время акта жевания.
Другими составными элементами периодонта являются: 1) кровеносные и лимфатические сосуды, 2) нервы и нервные рецепторы, 3) эпителиальные тельца Малассе, 4) иногда цемент (в виде цементиклей).
Сосуды в большинстве своем ориентированы параллельно длинной оси корня; их больше в задних, чем в передних, зубах, и в зубах нижней челюсти, чем верхней.
От них отходят капилляры, формирующие сплетение вокруг корня. Часть капилляров периодонта относится к фенестрированному типу, обладающему повышенной проницаемостью, что необходимо для обеспечения быстрого транспорта воды в основное вещество периодонта с целью компенсации давления в периодонтальной щели к жевательньм нагрузкам, воздействующим на зуб.
Вены, собирающие кровь из периодонта, направляются в одноименные артериям венозные коллекторы альвеолярного отростка верхней челюсти и альвеолярной части нижней челюсти.
Система лимфатических сосудов развита слабо. В ткани периодонта располагаются немногочисленные лимфатические капилляры, продолжающиеся в собирательные лимфатические сосуды, имеющие клапаны и идущие рядом с венами.
Периодонт получает как афферентную, так и эфферентную иннервацию. Афферентные нервы отходят к периодонту от зубных ветвей альвеолярных нервов (проходят в периодонте до десны) и от нервов, проникающих в отверстия межзубной и межкорневой костных перегородок (фолькмановские каналы) из верхнего и нижнего зубных сплетений, которые направляются в сторону верхушки корня или коронки.
Волокна из обоих источников, смешиваясь, образуют в периодонтальном пространстве нервное сплетение.
Нервные окончания периодонта относятся преимущественно к механорецепторам и болевым рецепторам (ноцицепторам). Наиболее богатой чувствительной иннервацией обладает периодонт у верхушки корня; она значительно меньше в области верхней трети корня и пучках циркулярной связки зуба.
Эфферентные нервы, представленные в основном симпатическими безмиелиновыми волокнами с окончаниями вокруг кровеносных сосудов, вероятно, участвуют в регуляции интенсивности кровотока в тканях периодонта. Парасимпатические волокна в перицементе не описаны.
Наличие в периодонте большого количества нервных волокон и чувствительных нервных окончаний (Л. И. Фалин, Б. Орбан и др.) определяет важную роль периодонта в рефлекторной регуляции жевательного давления, а также в качестве своеобразного органа осязания. Учитывая разнообразие форм рецепторов, заложенных в периодонте, можно предполагать, что их функция не ограничивается восприятием тактильных раздражений и жевательного давления. Несомненно, что раздражение рецепторов может явиться источником сильных болевых ощущений.
Нормальное функционирование периодонта, несомненно, связано с клетками, большое количество которых находится между волокнами периодонтальной связки. Эти клетки образуют и поддерживают все части прикрепляющего аппарата зуба. Среди них основными являются фибробласты, продуцирующие коллагеновые волокна периодонта, цементобласты (синтез цемента) и остеобласты (образование альвеолярной кости).
Преобладающим типом клеток периодонта являются фибробласты. Это уплощенные отростчатые клетки, лежащие вдоль коллагеновых волокон, которые образуют многочисленные соединения друг с другом посредством десмосом, щелевых и плотных соединений, формируя в периодонте единую трехмерную сеть. Клетки характеризуются значительным развитием гранулярной ЭПС и комплекса Гольджи, что свидетельствует об их активной синтетической функции.
Обнаружение во многих фибробластах лизосом, содержащих частично переваренные коллагеновые фибриллы, указывает на их фибропластические функции, т. е. направленные на разрушение межклеточного вещества периодонта. Основная функция этих клеток – образование и выделение коллагена.
Коллагеновые волокна периодонтальной связки подвергаются почти непрерывному процессу образования и перестройки (моделирования) в связи с функцией зуба, направлением жевательных усилий мышц, изменением окклюзионных соотношений зубных рядов и т. п. Оба этих вида деятельности выполняются, вероятно, системой фибробласт – фиброкласт одновременно.
Цементобласты образуют слой клеток периодонтальной поверхности цемента корня зуба. Это клетки вариабельной (чаще отростчатой) формы с округлым ядром и хорошо развитым синтетическим аппаратом. Они образуют в период активного формирования цемента непрерывный слой, вырабатывающий прецемент (необызвествленный органический матрикс цемента), который в дальнейшем подвергается минерализации.
Остеобласты располагаются в периодонте на поверхности стенки зубной альвеолы, формируя в активном состоянии непрерывный слой клеток, которые вырабатывают остеоид и осуществляют в дальнейшем его минерализацию.
В процессе возрастной перестройки костной ткани альвеолы остеобласты тесно взаимодействуют с остеокластами и одонтокластами. Последние появляются при резорбции (рассасывании) корней молочных зубов, а также при ряде патологических состояний, связанных с процессами в тканях корня.
В небольших количествах в интерстициальной соединительной ткани периодонта содержатся также макрофаги, лейкоциты (эозинофильные гранулоциты, моноциты, лимфоциты), тучные клетки. Они обеспечивают развитие и течение защитных реакций, поэтому при воспалительных процессах их содержание резко увеличивается.
Непосредственно около цемента часто обнаруживаются группы (островки) эпителиальных клеток – эпителиальные островки Малассе, которые образуются в период формирования корня зуба вследствие распада корневого эпителиального влагалища. Они более многочисленны сначала в апикальной части корня, а в более позднем возрасте – в шеечном отделе.
Наибольшее количество телец Малассе отмечается в детском возрасте (10–15 лет), в дальнейшем оно снижается. Микроскопически – это компактные скопления мелких клеток с относительно крупными ядрами и цитоплазмой, богатой гликогеном.
По морфологическим признакам выделяют три типа телец Малассе: 1) покоящиеся; 2) дегенерирующие; 3) пролиферирующие. В дегенерирующих островках присутствуют темные клетки с пикнотическими ядрами, подвергающиеся постепенному разрушению и обызвествлению с образованием кальцификатов. Пролиферирующие тельца имеют крупные размеры с клетками, характеризующимися заметной синтетической и пролиферативной активностью.
С возрастом отмечается увеличение телец Малассе 3‑го типа. Функция этих образований неясна. Эти клетки жизнеспособны, метаболически активны и, по некоторым представлениям, играют определенную роль в образовании кист периодонта, а также развития злокачественных опухолей.
Кроме описанных клеток, в периодонте находится также большое количество недифференцированных эктомезенхимальных клеток, происходящих из зубного фолликула (мешочка). Эти полипотентные клетки способны видоизменяться в любую клетку периодонта. Они являются основным резервом для всех клеток периодонтальной связки в течение жизни индивидуума.
В периодонте, как отмечалось, находится большое количество пучков коллагеновых волокон, натянутых между стенками зубной альвеолы челюсти и цементом корня зуба. Каждый пучок одним своим концом внедряется в кость альвеолярной стенки, другим – в цемент, причем их терминальные участки в обеих тканях называются прободающими (шарпеевскими) волокнами. Последние в кости обычно имеют значительно большую толщину (10–20 мкм), чем в цементе (3–5 мкм).
Особенностью коллагеновых фибрилл периодонта является их сравнительно небольшой диаметр – в среднем 50–60 нм, что в 2–4 раза меньше, чем диаметр таких фибрилл в сухожилии. Они характеризуются слегка волнообразным ходом, т. е. способны незначительно удлиняться при натяжении. В связи с этим, несмотря на отсутствие у них эластичности, коллагеновые волокна могут обеспечивать ограниченные движения зуба.
Промежутки между пучками плотной соединительной ткани заполнены прослойками рыхлой соединительной ткани, в которых проходят кровеносные и лимфатические сосуды, нервы, находятся специальные рецепторы, а также цементикли и тельца Малассе. Здесь же имеется небольшое количество тонких эластических волокон, по большей части связанными с кровеносными сосудами.
В последнее время в периодонте описан в дополнение к коллагеновым и эластическим волокнам 3‑й тип – окситалановые волокна диаметром 0,5–1,0 мкм, которые относят к преэластическим волокнам. Они многочисленны в области шейки зуба и располагаются обычно в центральном отделе периодонтального пространства аксиально, образуя вокруг корня трехмерную сеть, пронизывающую пучки коллагеновых волокон под прямым углом.
Пучки коллагеновых волокон в различных отделах периодонтального пространства имеют различное направление, обеспечивая прочную связь зуба с костной альвеолой. Так, у краев альвеолы они натянуты почти в горизонтальном направлении, образуя циркулярную связку зуба. Волокна этой связки одним своим концом укреплены в цементе вблизи цементоэмалевой границы, а другим вплетаются в соединительную ткань десны или прикрепляются к гребню альвеолярного отростка.
В составе циркулярной связки зуба выделяют транссептальные волокна периодонта, которые идут от цемента шейки одного зуба к такому же цементу смежного зуба в медиально‑периферическом направлении над гребешком альвеолярной кости в толще межзубного сосочка. Они образуют потенциально плотный барьер из соединительной ткани, задерживающий распространение инфекции, воспаления, вредных веществ из полости рта в периодонтальное пространство и к корню зуба.
Совокупность волокон периодонта, идущих в радиальном направлении в окружности шейки зуба и вершины корня, ограничивает возможность боковых (качательных) движений зуба при жевании.
У зубов, лишенных антагонистов и, следовательно, не функционирующих, в периодонтальной связке утрачивается правильное расположение пучков коллагеновых волокон и она становится очень тонкой [Кулидж Е., 1957].
Напротив, при повышенной нагрузке на зуб наблюдаются утолщение периодонтальных волокон и перестройка окружающей корень зуба альвеолярной кости, а также отложение новых слоев цемента на поверхности корня. Одновременно с этим в вещество цемента включаются новые волокна периодонта, возникшие благодаря деятельности фибробластов. Эти волокна приходят на смену старым дегенерировавшим волокнам перицемента. Этим объясняется, например, возможность перемещения зуба в процессе ортодонтического лечения без нарушения его связей с окружающими тканями [Орбан Б.,1984].
Оценивая в целом роль и значение пучков коллагеновых волокон периодонта, многие авторы рассматривают их как подвешивающий аппарат, который предохраняет сосудисто‑нервный пучок, вступающий в отверстие верхушки корня зуба, от сдавливания во время жевания.
Другие исследователи [Катц А., 1961; Быков В., 1996] считают основной ролью периодонта (помимо укрепления корня зуба в альвеоле) трансформацию жевательного давления и передачу его равномерно на все стенки костной зубной ячейки, а не только на ее пришеечную часть.
Возрастные изменения. В периодонте в течение всей жизни индивидуума постоянно происходят пластические процессы обновления его составных элементов: фибробластов и других клеток, коллагеновых волокон и межклеточного вещества.
Интенсивность обновления коллагена в периодонте в 2 раза выше, чем в десне, и в 4 раза выше, чем в коже. Высокая скорость обновления коллагена – свидетельство важной непрерывной перестройки периодонта в постоянно протекающих процессах адаптации поддерживающего аппарата зуба к меняющимся нагрузкам.
С возрастом уменьшаются пластические возможности клеток периодонта, что находит выражение в сокращении количества активных клеток (фибробласты, остеобласты, цементобласты), синтеза коллагеновых волокон, ухудшении кровоснабжения тканей перицемента. Отмечается уменьшение ширины периодонтальной щели. Эти изменения, несомненно, вызывают ослабление фиксирующего аппарата корня зуба, его барьерных свойств, увеличение подвижности зубов, что с неизбежностью приводит к необходимости врачебного вмешательства.
Глотка
Глотка (pharynx) – орган, где происходит перекрещивание дыхательного и пищеварительного путей. Состоит из 3 отделов:
1) носового (носоглотка), эпителий многорядный, призматический, реснитчатый, собственная пластинка содержит лимфоидные узелки;
2) ротового (ротоглотка), эпителий многослойный, плоский, неороговевающий;
3) гортанного, эпителий многослойный, плоский, неороговевающий.
Хорошо выражены все оболочки этого отдела пищеварительного тракта. В подслизистой основе носоглоточного отдела располагаются лимфоидная ткань трубных и глоточной миндалин и концевые отделы: смешанный: желез. В ротовом и гортанных отделах имеются слизистые железы и сеть эластических волокон.
Пищевод
Пищевод (oesophagus) – трубчатый орган, соединяющий глотку с желудком (рис. 14.1). Стенка образована 3 оболочками.

Рис. 14.1. Пищевод; поперечный срез. ×92.
1 – просвет пищевода; 2 – слизистая оболочка; 3 – многослойный плоский неороговевающий эпителий; 4 – собственная пластинка; 5 – мышечная пластинка; 6 – подслизистая основа; 7 – собственные железы; 8 – внутренний циркулярный слой мышечной оболочки; 9 – наружный продольный слой; 10 – адвентициальная оболочка.
Слизистая оболочка состоит из многослойного плоского неороговевающего эпителия, содержит интраэпителиальные лимфоциты и дендритные антигенпредставляющие клетки. Собственная пластинка слизистой оболочки – это рыхлая волокнистая соединительная ткань, вдающаяся в эпителий высокими сосочками. В нижней трети и в области перстневидного хряща располагаются кардиальные железы пищевода (простые трубчатые разветвленные), в концевых отделах которых находятся клетки, вырабатывающие муцины, париетальные клетки (вырабатывающие хлориды), эндокринные (ЕС– и ECL) клетки. Мышечная пластинка слизистой оболочки представлена одним продольным слоем гладко‑мышечных клеток. Подслизистая основа образована рыхлой волокнистой соединительной тканью, которая вместе с мышечной пластинкой формирует продольные складки. В ней располагаются лимфоидные узелки, подслизистое нервное сплетение, концевые отделы альвеолярно‑трубчатых собственных желез пищевода, выделяющих слизь, которая содержит бикарбонатные ионы и лизоцим.
Мышечная оболочка представлена 2 слоями: внутренним циркулярным и наружным продольным. В верхней трети пищевода мышечная оболочка состоит из поперечнополосатых мышечных волокон, в нижних двух третях – из гладкой мышечной ткани, на границе между ними – из их сочетания. Между слоями располагается межмышечное нервное сплетение.
Адвентициальиая оболочка образована рыхлой волокнистой соединительной тканью, в брюшном отделе – серозной оболочкой.
Желудок
Желудок (gaster) – объемистое мешковидное образование (рис. 14.2).

Рис. 14.2. Дно желудка. ×92.
1 – слизистая оболочка: 1а – железистый эпителий, 1б – собственная пластинка, 1в – мышечная пластинка; 2 – желудочные ямочки; 3 – собственные железы; 4 – подслизистая основа; 5 – мышечная оболочка: 5а – внутренний косой слой, 5б – средний циркулярный слой, 5в – наружный продольный слой; 6 – серозная оболочка.
Анатомически выщеляют 4 части: кардиальную, тело, дно и пилорическую; гистологически – три, так как тело и дно имеют сходное строение. Стенка образована тремя оболочками (см. рис. 14.2).
Слизистая оболочка имеет сложный рельеф: образует складки, поля, ограниченные бороздками и ямками (железы желудка). Состоит из 4 слоев:
а) эпителий однослойный призматический, одинаковый во всех отделах желудка, обновляется в течение 1–3 сут. Секретирует слизь, содержащую бикарбонаты, нейтрализующие соляную кислоту, образующую защитный барьер (барьер проницаем для алкоголя и аспирина);
б) собственная пластинка слизистой – это рыхлая волокнистая соединительная ткань, содержит много кровеносных и лимфатических сосудов, здесь же располагаются железы желудка (около 15 млн): простые, трубчатые, разветвленные собственные (фундальные), кардиальные и пилорические. В собственной пластинке имеются диффузные скопления лимфоидной ткани и лимфоидные узелки;
в) мышечная пластинка состоит из 3 слоев гладко‑мышечных клеток;
г) подслизистая основа образована рыхлой волокнистой соединительной тканью, содержит много эластических волокон, крупных сосудов, в ней располагается подслизистое нервное сплетение.
Мышечная оболочка состоит из 3 слоев гладких мышечных волокон: внутреннего слоя косых волокон, среднего – круговых (наиболее развит в области привратника, образуя пилорический сфинктер) и наружного – продольных волокон. Между слоями – прослойки соединительной ткани, а также межмышечное нервное сплетение.
Серозная оболочка образована мезотелием и подлежащей соединительной тканью (подсерозная основа).
Железы желудка. 1. Собственные, или фундальные, железы располагаются в теле и дне желудка. Самые многочисленные, образуют группы по 3–7, впадают в желудочные ямки. Железы трубчатые, слабо разветвленные, состоят из узкой шейки, удлиненного тела и дна. Различают 4 типа клеток: главные, париетальные, шеечные и эндокринные.
Главные клетки наиболее многочисленные в нижней части и дне желез, пирамидальные или цилиндрические. Клетки базофильны, хорошо развиты ЭПС, комплекс Гольджи, содержат в цитоплазме крупные апикально расположенные зимогенные гранулы (пепсиноген, проферменты).
Париетальные (обкладочные) клетки более многочисленны в верхней части желез. Они крупные, пирамидной формы, узкой апикальной частью вдаются между шеечными слизистыми и главными клетками. Цитоплазма оксифильна, ядро расположено в центре, много митохондрий с развитыми кристами. Имеют особые внутриклеточные секреторные канальцы в виде узких щелей со множеством микроворсинок. По периферии канальцев – комплекс мембранных пузырьков и трубочек (резерв мембраны, содержащей ионные насосы), сливающихся с канальцами при активной секреции. Через апикальный полюс секретируют ионы H и CL, которые, соединяясь, образуют HCL, создавая кислую среду в желудке (рН < 2,0), обеспечивающую разрушение белков, превращение пепсиногена в пепсин и угнетающую рост патогенных микроорганизмов. Через базальный полюс выделяют гидробикарбонаты, нейтрализованную HCL из просвета желудка. Синтезируют антианемический фактор, который в комплексе с витамином В12 всасывается в подвздошной кишке.
Слизистые шеечные клетки немногочисленны, располагаются в шейке железы поодиночке или группами. Цитоплазма слабобазофильна. Клетки часто делятся и рассматриваются как камбиальные клетки эпителия слизистой оболочки желудка. Выделяют слизь, предохраняющую стенку желудка от повреждения.
Эндокринные клетки располагаются в дне желез, они треугольные, овальные и полигональной формы. Ядро находится на апикальном полюсе, на базальном – плотные секреторные гранулы, ограниченные мембранами. Клетки окрашиваются солями серебра и хрома (хромаффинные гранулы), содержат пептидные гормоны и амины. Относятся к APUD‑системе. Различают EC‑клетки – вырабатывают серотонин, мотилин, вещество Р, усиливают моторику кишечника; ECL‑клетки – выделяют гистамин, усиливающий секрецию HCL в желудке, и G‑клетки – выделяют гастрин, активизирующий секрецию HCL и пепсиногена железами желудка.
2. Кардиальные железы трубчатые, разветвленные, концевые отделы содержат слизистые клетки со светлой цитоплазмой, вырабатывают мукоидный секрет, бикарбонаты, хлориды калия и натрия. В железах встречаются главные, париетальные и эндокринные клетки.
3. Пилорические железы трубчатые, с сильно разветвленными и извитыми концевыми отделами, располагаются в пилорическом отделе желудка, образованы слизистыми, париетальными и эндокринными (G и EC) клетками.
Тонкая кишка
Тонкая кишка (intestinum tenue) состоит из 3 отделов: двенадцатиперстной (рис. 14.3), тощей (рис. 14.4) и подвздошной кишок, которые имеют сходное строение. Общая длина их 6–7 м посмертно, прижизненно 3 м. Стенка образована тремя оболочками.

Рис. 14.3. Двенадцатиперстная кишка. ×120.
1– слизистая оболочка (ворсинка): 1а – однослойный призматический (каемчатый) эпителий, 1б – бокаловидные клетки, 1в – собственная пластинка, 1 г – крипта, 1д – мышечная пластинка; 2 – подслизистая основа: 2а – дуоденальные железы; 3 – мышечная оболочка: 3а – внутренний циркулярный слой, 3б – наружный продольный слой; 4 – адвентициальная оболочка.
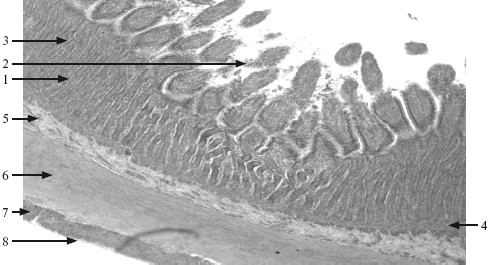
Рис. 14.4. Тощая кишка. ×15.
1 – слизистая оболочка; 2 – эпителий; 3 – собственная пластинка с криптами; 4 – мышечная пластинка слизистой оболочки; 5 – подслизистая основа; 6 – внутренний циркулярный слой мышечной оболочки; 7 – наружный продольный слой мышечной оболочки; 8 – серозная оболочка.
Слизистая оболочка – однослойный призматический каемчатый эпителий, собственная пластинка, мышечная пластинка и подслизистая основа. Рельеф оболочки сложный:
Циркулярные складки (складки Керкринга) сформированы всеми слоями слизистой оболочки, они занимают от 1/2 до 2/3окружности стенки кишки; их около 800, наиболее развиты в тощей кишке;
Кишечные ворсинки – пальцевидные выпячивания слизистой оболочки, образованные собственной пластинкой. Покрыты призматическим каемчатым эпителием. Ворсинки двенадцатиперстной кишки шире и короче, листовидные. В дистальном направлении становятся выше и уже. Общее количество ворсинок тонкой кишки достигает 4–5 млн;
Кишечные крипты (железы) – трубчатые углубления эпителия в собственную пластинку; открываются в пространство между ворсинками. Общее количество крипт более 150 млн. Содержат камбиальные элементы. Обеспечивают выработку пищеварительных соков и слизи.
Эпителий содержит 5 типов клеток: каемчатые, бокаловидные, клетки с ацидофильными гранулами (клетки Панета), недифференцированные или малодифференцированные (бескаемчатые), эндокринные. В пейеровых бляшках имеются специализированные М‑клетки.
Каемчатые клетки составляют основную массу эпителия ворсинок и верхушки крипт, имеют высокопризматическую форму, хорошо развитые органеллы; продолжительность жизни 1,5–3 сут. На апикальной поверхности располагаются микроворсинки (до 3000), покрыты гликокаликсом, образуют щеточную каемку, содержащую ферменты, обеспечивающие пристеночное пищеварение, расщепление олигомеров до мономеров, всасываемых клеткой. Здесь осуществляется переваривание белков и углеводов до аминокислот и моносахаров. Энтероциты способны транспортировать неизмененные макромолекулы (фактор роста, иммуноглобулины), что имеет особое значение для детей грудного возраста. Жиры эмульгируются и расщепляются до свободных жирных кислот и моноглицеридов, при этом образующиеся хиломикроны (окруженные мембраной гликолипопротеины) поступают в лимфатический капилляр ворсинки (млечные пути). Осуществляют всасывание воды и солей.
Бокаловидные клетки – одноклеточные слизистые железы, располагаются среди каемчатых клеток в ворсинках и криптах. Продолжительность жизни 2–4 сут. Секреторный цикл активен, выделяют слизь.
Клетки Панета (с ацидофильными гранулами) пирамидной формы, в узкой апикальной части их располагаются крупные ацидофильные гранулы с белково‑полисахаридным комплексом, содержат цинк, лизоцим (бактерицидный фермент). Располагаются в дне крипт, обновляются медленно (3–4 нед).
Недифференцированные (бескаемчатые) клетки находятся в глубине крипт, являются источником обновления эпителия крипт и ворсинок. Узкие, призматические клетки, стволовые, активно делящиеся.
Эндокринные клетки многочисленны в криптах. Наиболее распространены: EC‑клетки, выделяющие серотонин, усиливающий моторику кишечника, мотилин, вещество Р; G‑клетки – гастрин, активирующий секрецию HCL и пепсиногена в желудке; I‑клетки – холецистокинин/панкреозимин, который повышает секрецию ферментов поджелудочной железой, усиливает сокращение желчного пузыря и желчеотделение; S‑клетки – секретин, усиливающий секрецию бикарбонатов поджелудочной железой; К‑клетки – ЖИП (желудочный ингибирующий пептид), понижающий секрецию и моторику желудка и секрецию инсулина; D‑клетки – соматостатин, угнетающий секрецию ГЭП‑клеток (ГЭП – гастроэнтеропанкреатические) и желез желудка. Интра‑эпителиальные лимфоциты располагаются между энтероцитами, преобладают Т‑лимфоциты (супрессоры, клетки памяти и киллеры).
Собственная пластинка представлена рыхлой волокнистой соединительной тканью, в которой располагается большое количество лимфоцитов (Т‑хелперы, плазматические клетки), эозинофилы, макрофаги, тучные клетки, система кровеносных сосудов, широкие лимфатические капилляры, гладко‑мышечные клетки на оси ворсинки, своим сокращением способствующие продвижению лимфы. Имеются солитарные и сгруппированные лимфоидные узелки (пейеровы бляшки), содержащие Т– и В‑лимфоциты.
В области пейеровой бляшки крипты и ворсинки мелкие или отсутствуют. В эпителии образуется купол пейеровой бляшки, встречаются М‑клетки (микроскладчатые), они ниже каемчатых, апикальный полюс покрыт немногочисленными микроворсинками со слабо развитым гликокаликсом. В «карманах» М‑клеток находятся лимфоциты, в бляшках – В‑лимфоциты, плазмоциты, Т‑лимфоциты, макрофаги, антигенпредставляющие дендритные клетки.
Мышечная пластинка слизистой оболочки образована 2 слоями гладко‑мышечных клеток.
В подслизистой основе встречаются лимфоидные узелки, элементы подслизистого нервного, венозного и лимфатического сплетений, в двенадцатиперстной кишке – железы (дуоденальные, или бруннеровы), сложные трубчатые, в секреторных клетках их концевых отделов содержатся гликопротеиды, а выводные протоки открываются в крипты. Секрет содержит слизь, обладающую щелочной реакцией, лизоцим, урогастрон (стимулирующий деление эпителиальных клеток и угнетающий секрецию соляной кислоты в желудке) и ферменты – дипепсидазу, амилазу, энтерокиназу.
В мышечной оболочке между двумя слоями (внутренним круговым и наружным продольным) располагается межмышечное нервное сплетение.
Серозная оболочка образована мезотелием и подлежащей соединительной тканью (подсерозной основой), в двенадцатиперстной кишке оболочка адвентициальная.
Толстая кишка
Толстая кишка (intestinum crassum) состоит из 4 отделов: слепой кишки с червеобразным отростком, ободочной кишки (включает восходящую, поперечную, нисходящую и сигмовидную) и прямой кишки.
Стенка кишки образована тремя оболочками: слизистой, мышечной и серозной (или адвентициальной) (рис. 14.5).

Рис. 14.5. Толстая кишка. х 140.
1 – слизистая оболочка; 2 – эпителий (каемчатые эпителиоциты и бокаловидные клетки); 3 – собственная пластинка; 4 – крипты; 5 – мышечная пластинка слизистой оболочки; 6 – подслизистая основа; 7 – мышечная оболочка (внутренний циркулярный слой).
Слизистая оболочка включает 4 слоя: эпителий, собственную пластинку и мышечную пластинку слизистой, подслизистую основу. Ворсинки отсутствуют, имеются полулунные складки, кишечные крипты глубже и располагаются чаще.
Эпителий однослойный призматический, состоит из призматических, бокаловидных, малодифференцированных и эндокринных клеток:
Призматические клетки высокие, каемка выражена слабо, вырабатывают и секретируют гликопротеины, обеспечивают процесс всасывания;
Бокаловидные клетки увеличиваются в количестве в дистальном направлении, выделяют слизь;
Малодифференцированные клетки лежат в глубине крипт, являются камбиальными. Обновление эпителия осуществляется в течение 6 сут;
Эндокринные клетки располагаются в дне крипт, встречаются ЕС– и ECL‑клетки. В рыхлой волокнистой соединительной ткани собственной пластинки находятся капилляры, нервные волокна, одиночные лимфоидные узелки, которые могут проникать в мышечную пластинку и подслизистую основу.
Мышечная пластинка состоит из 2 слоев гладко‑мышечных клеток.
Подслизистая основа содержит большое количество эластичных волокон и жировой ткани, подслизистые нервное, венозное и лимфоидное скопления.
Мышечная оболочка образована 2 слоями гладкой мышечной ткани: внутренним круговым и наружным продольным в виде 3 лент; за счет их меньшей длины образуются мешковилные выпячивания (гаустры). Между слоями в прослойке соединительной ткани расположено межмышечное нервное сплетение.
Серозная оболочка покрывает полностью стенки органа (слепая, ободочная кишки) или замещается частично адвентициальной (восходящая и нисходящая кишки).
Червеобразный отросток – пальцевидный вырост слепой кишки с узким звездчатым или треугольным (у детей) просветом, который содержит клеточный детрит; способен облитерироваться. Стенка толстая из‑за наличия в собственной пластинке и подслизистой основе многочисленных лимфоидных фолликулов. Состоит из 3 оболочек, как и стенка толстой кишки.
Червеобразный отросток выполняет защитную функцию, является периферическим органом иммунной системы. Обеспечивает поглощение антигенов из просвета толстой кишки. Содержит эффекторные В– и Т‑лимфоциты. Воспаление червеобразного отростка (аппендицит) может вызвать разрушение и разрыв его стенки и воспаление брюшины – перитонит.
Прямая кишка – дистальный участок толстой кишки, завершается заднепроходным (анальным) каналом.
Слизистая оболочка заднепроходного канала образует 5–10 продольных складок – заднепроходных (анальных) складок (морганьевы столбы), соединенные поперечными складками. Между столбами образуются заднепроходные синусы. Крипты в дистальном направлении укорачиваются и исчезают. Эпителий однослойный призматический замещается в заднепроходном канале многослойным плоским неороговевающим, в переходной зоне – многослойным призматическим или кубическим, в области наружного сфинктера заднего прохода сменяется кожным многослойным плоским ороговевающим эпителием, появляются сальные, потовые железы и волосы.
Анальные железы – рудиментарные образования, располагаются в подслизистой основе и открываются в заднепроходные синусы, выстланы призматическим эпителием, вырабатывающим слизь, окружены лимфоидной тканью. В подслизистой основе располагается геморроидальное венозное сплетение, состоящее из 2 отделов: внутреннего (в подслизистой основе заднепроходных столбов) и наружного (в области наружного сфинктера). Варикозное расширение этих вен является заболеванием – геморроем. Внутренний сфинктер заднего прохода представлен циркулярно расположенным слоем гладко‑мышечных клеток, наружный образован поперечнополосатой мышечной тканью.
Дата добавления: 2019-02-12; просмотров: 467; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!