Морфологическая классификация нервных окончаний
Свободные нервные окончания:
– терминали дендрита располагаются непосредственно между клетками иннервируемой ткани и не окружены глиоцитами;
– характерная локализация – эпителиальная ткань (в основном многослойный плоский эпителий);
– не обладают строгой специфичностью восприятия раздражений. Несвободные нервные окончания:
– образованы разветвлением терминали дендрита и расположенными вокруг глиоцитами. Такие нервные окончания называют несвободными некапсулированными;
– к ним относят осязательные мениски, или диски Меркеля, являющиеся специализированными эпителиальными рецепторными окончаниями (механорецепторы);
– несвободные нервные окончания могут сопровождаться более или менее тонким соединительно‑тканным слоем в виде капсулы. Такие нервные окончания называют несвободными капсулированными;
– капсулированные чувствительные нервные окончания наблюдаются в соединительной и мышечной тканях;
– в соединительной ткани располагаются пластинчатые тельца, или тельца Фатера – Пачини; луковицеобразные тельца, или тельца Гольджи – Маццони; осязательные тельца, или тельца Мейснера; концевые колбы Краузе; тельца Руффани; генитальные тельца, или тельца Догеля, а также капсулированные тельца птиц – тельца Хербста и тельца Грандри;
– к капсулированным рецепторам мышечной ткани относятся нервно‑мышечные веретена;
|
|
|
– в теле человека наиболее распространенными являются тельца Фатера – Пачини, являющиеся механорецепторами. Они располагаются около сухожилий и суставов, в молочной железе, коже, брыжейке тонкой кишки, поджелудочной железе, соединительной ткани внутренних органов и около кровеносных сосудов;
– они овальные, состоят из дендрита чувствительного нейрона, соединительно‑тканной капсулы и нейроглиальных клеток, расположенных между ними;
– дендрит вместе с нейроглиальными клетками образует внутреннюю колбу тельца;
– внутренняя колба, в свою очередь, покрыта соединительно‑тканной оболочкой, называемой наружной колбой, образованной уплощенными фибробластоподобными клетками, которые концентрически наслаиваются друг на друга;
– в сосочковом слое кожи расположены овальные осязательные тельца Мейснера, которые являются механорецепторами. Дендрит чувствительного нейрона разветвляется в тельце в виде плоской спирали, вокруг которой располагаются многочисленные нейроглиальные клетки, тесно окружающие дендрит и образующие вместе с ним внутреннюю колбу тельца. Наружная колба образована соединительно‑тканной капсулой, богатой эластическими волокнами;
|
|
|
– к механорецепторам подобного строения относятся тельца Гольджи – Маццони, располагающиеся в сухожилиях;
– весьма распространенными в теле человека являются концевые колбы Краузе, которые являются терморецепторами, располагаются в коже, слизистых и серозных оболочках;
– температурные раздражения воспринимают также капсулированные тельца Руффини, локализованные там же, где и колбы Краузе;
– генитальные тельца Догеля, в большом количестве располагающиеся в соединительной ткани половых органов (клитор, головка полового члена и т. д.), по строению напоминают колбы Краузе, но отличаются от них тем, что в их образовании участвует несколько дендритов чувствительных нейронов; вокруг их капсулы располагается множество мелких кровеносных сосудов;
– генитальные тельца являются механо– и барорецепторами, поскольку реагируют на изменения кровяного давления.
Двигательные (эфферентные) нервные окончания образуются аксонами мотонейронов на рабочих органах. Выделяют нервно‑мышечное и нейросекреторное окончания.
Миелиновое волокно подходит к поверхности скелетного мышечного волокна и в месте их контакта формируется так называемое нервно‑мышечное окончание, или нервно‑мышечный синапс (моторная бляшка). Принцип его морфофункциональной организации аналогичен таковому межнейрального химического синапса. Между поверхностью аксона и сарколеммой мышечного волокна имеется синаптическое пространство;
|
|
|
– различают первичное и вторичное синаптические пространства, или субнейральный аппарат;
– первое располагается между сарколеммой и аксолеммой, примыкая к последней, а второе образовано глубокими складками сарколеммы, переходящими в Т‑систему мышечного волокна;
– базальная мембрана мышечного волокна переходит на нервное волокно. Сарколемма формирует многочисленные складки. Саркоплазма теряет характерную поперечную исчерченность. Окончание аксона содержит многочисленные митохондрии и синаптические пузырьки с ацетилхолином. То же самое можно сказать и по отношению к саркоплазме мышечного волокна, однако в ней нет синаптических пузырьков, отсутствуют миофибриллы, но много митохондрий.
Двигательные нервные окончания в гладких мышцах отличаются от моторных бляшек тем, что аксон мотонейрона, ветвясь, образует многочисленные колбовидные конечные расширения, которые обращены к группам гладких миоцитов, но находятся от последних на некотором удалении (синаптическое пространство может достигать 80–100 нм).
|
|
|
При формировании нейросекреторного окончания аксон также заканчивается на некотором расстоянии от железистых клеток. Выделяемый нейромедиатор диффузно достигает группы секреторных клеток железы и воздействует на нее.
Тесты и вопросы для самоконтроля
Выберите один или несколько правильных ответов:
1. Производные нервной трубки:
а) пирамидные нейроны коры больших полушарий головного мозга;
б) псевдоуниполярные нейроны спинномозговых узлов;
в) эпендимоциты;
г) меланоциты;
2. По своему строению нейрон отличается от других клеток организма:
а) наличием отростков;
б) наличием контактов между клетками;
в) наличием полярных отростков и синапсов;
г) наличием одного диплоидного ядра.
3. Какие специфические структуры характерны для нервной клетки:
а) лизосомы и комплекс Гольджи;
б) базофильное вещество;
в) митохондрии;
г) фибриллярные структуры.
4. Структура периферического нерва:
а) эндоневрий;
б) кровеносные сосуды;
в) фибробласты;
г) олигодендриты;
д) миоциты.
5. В миелиновом нервном волокне различают:
а) перехват Ранвье;
б) насечки Шмидта – Лантерманна;
в) мезаксон;
г) протоплазматические астроциты.
6. Эпендимная глия:
а) выстилает центральный канал спинного мозга;
б) имеет реснички;
в) секретирует цереброспинальную жидкость;
г) выполняет функцию иммунной защиты.
7. Для клеток микроглии характерно:
а) мезенхимное происхождение;
б) участие в иммунном ответе в ЦНС;
в) способность к обновлению;
г) многочисленные лизосомы.
8. Периневрий:
а) тонкий слой соединительной ткани вокруг каждого нервного волокна;
б) содержит клетки, связанные плотными контактами;
в) ретикулярная ткань вокруг пучков нервных волокон;
г) контролирует проницаемость и поддерживает гомеостаз эндоневрия.
9. По аксону транспортируется все, кроме:
а) белковых молекул;
б) везикул;
в) нейромедиаторов;
г) митохондрий;
д) рибосом.
10. К механорецепторам относятся:
а) тельца Руффини;
б) тельца Догеля;
в) тельца Мейсснера;
г) колбы Краузе;
д) тельца Фатера – Пачини.
Ответы
1: а, в.
2: в.
3: б.
4: а, б, в, г.
5: а, б, в.
6: а, б, в.
7: а, б, в.
8: б, г.
9: д.
10: б, в, д.
Глава 9
Нервная система
Нервная система – система органов, построенных из нервной ткани. Многие из этих органов имеют чрезвычайно сложное строение. В нервной системе выделяют центральный (спинной и головной мозг) и периферический (нервы, узлы, сплетения) отделы. Существует также подразделение на анимальный (соматический) и автономный (вегетативный) отделы.
Центральная нервная система
Центральная нервная система (ЦНС) – это наиболее сложная в структурном и функциональном отношении структура организма. Органы ее представляют собой сочетание хорошо отличающихся по окраске, даже в нативном состоянии, серого и белого веществ. В разных отделах ЦНС количество и взаимное расположение серого и белого веществ различны.
Серое вещество по своему гистологическому строению является сочетанием огромного количества мультиполярных нейроцитов разных величины и формы. Обязательными компонентами серого вещества являются также нервные волокна и глиоциты. По приблизительным подсчетам ЦНС содержит в 10 раз больше клеток глии, чем нейроцитов.
Белое вещество образовано мякотными нервными волокнами и расположенными между ними клетками глии.
В единице объема серого вещества значительно больше кровеносных капилляров, чем в белом веществе.
Спинной мозг. В спинном мозге хорошо различимы центрально расположенное серое вещество и окружающее его белое вещество (рис. 9.1).
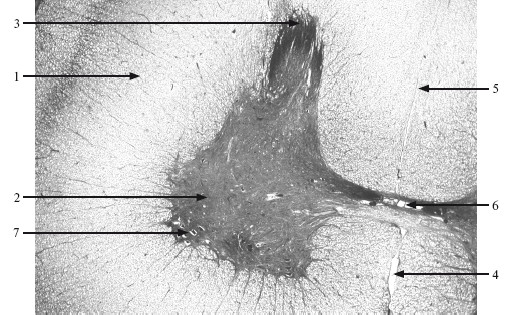
Рис. 9.1. Спинной мозг. ×56.
1 – белое вещество (боковой столб); 2 – серое вещество (передний рог); 3 – задний рог; 4 – передняя срединная щель; 5 – задняя спайка; 6 – центральный канал; 7 – двигательные нейроны.
Соотношение серого и белого вещества неодинаково в разных отделах спинного мозга. Серое вещество в максимальной степени развито в шейном и поясничном утолщениях, а масса белого вещества постепенно нарастает от каудального отдела к краниальному.
Серое вещество в поперечном сечении имеет вид бабочки или буквы Н. Средняя часть серого вещества – серая спайка, в которой проходит центральный канал. Серое вещество образует резко ограниченные выступы, называемые рогами. Различают передние, задние и боковые (в нижних шейных, грудных и верхних поясничных сегментах спинного мозга) рога.
Наиболее важными в функциональном отношении структурами серого вещества являются многоотростчатые нервные клетки. Скопления этих клеток, характеризующихся сходными размерами, структурой и функцией, образуют ядра, являющиеся по существу нервными центрами спинного мозга. Помимо клеток, в сером веществе имеется огромное количество нервных волокон – отростков нейроцитов, а также множество нейроглиальных клеток и соединительная ткань с сосудами.
Нейроглиальные элементы серого вещества представлены протоплазматическими и волокнистыми астроцитами, клетками олигодендроглии, а также эпендимой, выстилающей центральный канал спинного мозга. Отростки эпендимоцитов вместе с другими элементами глии образуют остов серого вещества.
Мультиполярные нейроциты серого вещества в зависимости от длины и расположения их нейритов подразделяются на три типа: корешковые, пучковые и внутренние.
Корешковые клетки имеют нейриты, выходящие из спинного мозга в составе передних корешков. Нейриты пучковых клеток проходят в пучки белого вещества (проводящие пути), а нейриты внутренних клеток не выходят за пределы спинного мозга. Проходя в белом веществе (несколько сегментов вверх и вниз), нейриты внутренних клеток снова входят в серое вещество и заканчиваются на его нейроцитах. Те клетки, отростки который выходят в одноименную половину белого вещества, называются гомомерными, а те, которые посылают свои нейриты на противоположную сторону, – гетеромерными.
Нейроны серого вещества образуют скопления – нервные пластинки; в заднем роге их шесть (I–VI), в боковом – одна (VII), в переднем роге – две (VIII и IX) и в центральном сером веществе одна (X). Нейроны пластинок формируют скопления – ядра.
Различают следующие ядра серого вещества спинного мозга.
В передних рогах расположены двигательные ядра. У человека в связи со сложной деятельностью верхних конечностей эти ядра особенно выражены в шейном утолщении. Они образованы наиболее крупными корешковыми нейроцитами спинного мозга, диаметр которых доходит до 11 мкм и более. Эти клетки имеют короткие разветвленные дендриты и длинные нейриты. Нейроны переднего рога формируют следующие ядра:
1) переднемедиальное;
2) заднемедиальное;
3) центральное и рядом с ним ядро диафрагмального нерва (С3–С5);
4) переднелатеральное и рядом с ним ядро добавочного нерва (С1–С6);
5) заднелатеральное;
6) зазаднелатеральное.
Аксоны нейронов двигательных ядер покидают спинной мозг в составе передних корешков, основную массу которых они составляют, и заканчиваются в скелетной мускулатуре. Мотонейроны медиальных ядер иннервируют мышцы туловища, латеральных – мышцы конечностей. Диафрагмальное ядро осуществляет двигательную иннервацию диафрагмы, а ядро добавочного нерва – грудино‑ключично‑сосцевидной и трапециевидной мышц. Среди клеток двигательных ядер различают α‑клетки, нейриты которых заканчиваются моторными окончаниями, и γ‑клетки, длинные отростки которых идут к мышечным веретенам.
γ‑Клетки не вызывают двигательного эффекта, хотя их аксоны (γ‑эфферентные волокна) заканчиваются эффекторными окончаниями. Обусловливая натяжение мышечных веретен, они видоизменяют деятельность рецепторов мышц, усиливая их афферентную импульсацию.
Двигательные клетки переднего рога посылают коллатерали своих аксонов к особой группе клеток, описанных Реншоу (вставочные нейроны), нейриты которых образуют синапсы на мотонейронах и тормозят их деятельность по принципу отрицательной обратной связи.
В заднем роге выделяют различные по своему микроскопическому строению части:
а) краевую зону Лиссауэра (I нервная пластинка), в которой располагается краевое ядро;
б) студенистое вещество Рональда (II нервная пластинка), в нем располагаются мелкие нейроны и много глиальных клеток;
в) собственное ядро заднего рога (III–VI пластины).
В боковом роге, у основания заднего рога, локализуется заднее грудное ядро (ядро Кларка).
В краевом ядре и студенистом веществе имеются мелкие нейроциты, но основными структурами также являются клетки глии (глиоциты).
Собственное ядро (n. proprius) заднего рога расположено у его основания и образовано гетеромерными пучковыми клетками. Нейриты этих клеток формируют в противоположной половине спинного мозга пучки белого вещества, осуществляющие связь спинного мозга с мозжечком и зрительными буграми (передний спиномозжечковый путь Говерса и передний и латеральный спиноталамические пути).
Кроме того, в задних рогах много диффузно расположенных мелких мультиполярных нейроцитов, являющихся внутренними ассоциативными клетками.
Задние грудные ядра или грудные столбы, расположенные на уровне C8–L2 боковых рогов спинного мозга, состоят из гомомерных пучковых клеток, нейриты которых в одноименной половине белого вещества образуют пучки (задний спиномозжечковый путь, или путь Флексига) и заканчиваются в мозжечке.
Автономные ядра боковых рогов выражены в тораколюм‑бальном (Th1–L2) и сакральном отделах спинного мозга. Выделяют ядра: а) промежуточно‑медиальное (nucl. intermediomedialis), к которому подходят афферентные пути от внутренних органов; б) промежуточно‑латеральное (nucl. intermediolateralis), содержащее преганглионарные нейроны вегетативной нервной системы; в) крестцовые парасимпатические ядра (S2–S4; nucll. parasympathici sacrales), иннервирующие органы малого таза.
Нейриты гомомерных корешковых клеток этих ядер выходят из спинного мозга в передних корешках и направляются к автономным нервным узлам.
Собственные (ассоциативные) ядра спинного мозга состоят из внутренних гомо– и гетеромерных клеток, рассеянных диффузно, без определенной группировки, преимущественно в промежуточной зоне серого вещества. Нейриты этих клеток за пределы спинного мозга не выходят и образуют собственные основные, или ассоциативные, пучки белого вещества.
Белое вещество спинного мозга состоит из продольно идущих мякотных нервных волокон. Опорной тканью белого вещества является нейроглия, преимущественно волокнистые астроциты. Белое вещество разделяется передними и задними корешками на три канатика: передний, задний и боковой.
Пучки нервных волокон белого вещества в канатиках образуют короткие пути, называемые собственными путями, или собственным аппаратом, спинного мозга, а также длинные пути, связывающие спинной мозг с различными отделами головного мозга.
Длинные пути разделяются на восходящие и нисходящие.
Восходящие пути. 1. Тонкий (пучок Голля) и клиновидный (пучок Бурдаха) пучки расположены в задних канатиках белого вещества. Эти пучки образованы мякотными нервными волокнами, которые являются нейритами псевдоуниполярных клеток спинномозговых узлов; по задним корешкам они входят в спинной мозг. В краевой зоне заднего рога эти волокна Т‑образно делятся на две ветви. Одна из них заканчивается синапсами на нейронах серого вещества, другая в составе тонкого и клиновидного пучков поднимается в продолговатый мозг и заканчивается здесь на собственных ядрах – ядре тонкого и ядре клиновидного пучков.
В медиально расположенном тонком пучке проходят нервные волокна от нижней половины тела, в латеральном клиновидном пучке – волокна, идущие от верхней части туловища. Эти пучки проводят глубокую проприоцептивную чувствительность (мышечно‑суставное чувство) и частично кожную.
2. Задний спиномозжечковый пучок (пучок Флексига) формируют волокна, берущие начало от гомомерных пучковых клеток заднего грудного ядра бокового рога. Эти волокна располагаются в дорсолатеральном отделе бокового канатика одноименной половины спинного мозга и направляются в кору червя мозжечка.
3. Передний спиномозжечковый пучок (пучок Говерса) расположен в вентролатеральном отделе бокового канатика белого вещества. Волокна этого пучка берут начало от гетеромерных пучковых клеток собственного ядра серого вещества заднего рога. Нейриты клеток данного ядра через серую спайку переходят на противоположную половину спинного мозга и в составе бокового канатика поднимаются вверх, заканчиваясь в коре червя мозжечка.
Оба спиномозжечковых пучка передают в мозжечок импульсы, возникающие в проприорецепторах, участвуют в регуляции сложных двигательных актов и поддержании равновесия тела.
4. Спиноталамические пучки – боковой и передний – осуществляют непосредственную связь спинного мозга с таламусом. Волокна их представляют собой аксоны гетеромерных пучковых клеток собственного ядра заднего рога. Расположены спиноталамические пучки в боковых и передних канатиках белого вещества. Эти пучки являются проводниками болевой и температурной (боковой пучок), а также тактильной (передний пучок) чувствительности. Помимо этих наиболее крупных восходящих путей, в белом веществе спинного мозга имеются и более мелкие: спинопокрышечный, идущий к четверохолмию, восходящий пучок переднего канатика, спинооливный и др.
Нисходящие пути. Эти пути спинного мозга могут быть разделены на две системы: пирамидную и экстрапирамидную.
Пирамидные пути (корково‑спинномозговые) берут начало от крупных пирамидных клеток коры больших полушарий. Нейриты этих клеток (до 1 000 000) в составе одного пучка доходят до границы между продолговатым и спинным мозгом. Здесь, в области так называемого перекреста пирамид, большая часть волокон переходит на противоположную сторону, т. е. образует перекрест, и составляет значительный участок бокового канатика. Другие волокна проходят, не перекрещиваясь, в передних канатиках белого вещества до ядер переднего рога своей и противоположной сторон. Таким образом возникает два корково‑спинальных пути: боковой – перекрещенный и передний – неперекрещенный (прямой). Оба заканчиваются на нейронах двигательных ядер переднего рога серого вещества разных сегментов спинного мозга. По этим путям проходят импульсы для волевых сознательных движений (целенаправленные рабочие движения). Лучше всего эти пути развиты у человека.
Экстрапирамидные пути берут начало от нейроцитов стволовой части головного мозга.
1. Красноядерно‑спинномозговой путь сформирован из нейритов нервных клеток красных ядер среднего мозга. По выходе из ядер волокна переходят на противоположную сторону (совершают перекрест) и располагаются в боковом канатике спинного мозга вентральнее бокового пирамидного пути; заканчиваются на двигательных ядрах переднего рога спинного мозга. У животных этот путь развит лучше, чем у человека, он проводит импульсы для непроизвольных автоматических движений, имеет значение в координации этих движений и регуляции тонуса мышц.
2. Покрышечно‑спинномозговой путь начинается от верхних бугорков четверохолмия, занимает медиальный отдел передних канатиков спинного мозга и заканчивается на двигательных клетках передних рогов спинного мозга. В четверохолмии заканчиваются волокна зрительного тракта и от затылочной области коры, поэтому идущие по этому пучку импульсы обеспечивают уточнение и направленность двигательных актов, связанных с сочетанным движением головы и глаз.
3. Латеральный и медиальный преддверно‑спинномозговые пути берут начало соответственно от латерального и медиального вестибулярных ядер продолговатого мозга, проходят в переднем канатике и заканчиваются на нейроцитах переднего рога спинного мозга. Эти пути имеют большое значение для правильной ориентировки тела в пространстве, так как в вестибулярных ядрах оканчиваются нейриты чувствительных клеток вестибулярного ганглия, осуществляющего восприятие раздражений от органа статического равновесия внутреннего уха.
Помимо перечисленных, имеются еще более мелкие пучки: ретикулоспинномозговые, задний продольный пучок, оливоспинномозговой и др.
Короткие пучки белого вещества спинного мозга проходят небольшие расстояния. Они располагаются во всех трех канатиках спинного мозга: передних, задних, боковых, окружая серое вещество. Это собственные (ассоциативные) пучки, связывающие отдельные сегменты спинного мозга, которые образованы нейритами гомо– и гетеромерных клеток собственного ядра заднего рога. Выходя в белое вещество, они проходят в нем на протяжении нескольких сегментов вверх или вниз, а затем снова входят в серое вещество и заканчиваются на мультиполярных нейронах двигательных ядер переднего рога.
Ствол головного мозга. В стволе мозга серое вещество образует ядра, содержащие в своем составе моторные, чувствительные и ассоциативные нейроны. Мотонейроциты посылают свои аксоны – двигательные волокна черепных нервов – на периферию. На чувствительных нервных клетках оканчиваются нейриты клеток чувствительных узлов головы и тела.
Ассоциативные нейроциты образуют ядра, на которых происходит переключение нервных импульсов, идущих из одного отдела ЦНС в другой (например, из спинного мозга и ствола в кору больших полушарий и наоборот). В силу важности и многообразия выполняемых функций следует остановиться на особенностях морфологии таких отделов ЦНС, как ретикулярная формация (formatio reticularis), области таламуса и гипоталамуса.
Ретикулярная формация (сетчатое образование) объединяет совокупность определенных участков ЦНС, располагающихся в виде обособленного тяжа в спинном, продолговатом и промежуточном мозге. Как самостоятельная субстанция она была описана еще в 1855 г. Ленхошеком. В. М. Бехтерев выделил в ней обособленные клеточные скопления – ядра.
Ядра ретикулярной формации в продолговатом мозге формируют дыхательный центр, описанный Н. А. Миславским. Они посылают импульсы в спинной мозг по ретикулоспинномозговому пути. В дне IV желудочка на уровне олив имеется другой участок этого образования, называемый сосудодвигательным центром, который был описан Ф. В. Овсянниковым. Выделены также центры глотания, защитных рефлексов (кашель, рвота, чиханье) и др.
При гистологическом исследовании в ретикулярной формации обнаружено большое количество нервных волокон, идущих в разных направлениях (отсюда название «сетчатая формация», данное Дейтерсом). В дорсомедиальном его отделе имеются преимущественно крупные мультиполярные нейроциты, аксоны которых, идущие в выше– и нижерасположенные отделы мозга, отдают множество коллатералей. Благодаря этому один нейроцит ретикулярной формации может иметь связи более чем с 27 000 других нейроцитов. В латеральном отделе преобладают мелкие чувствительные нейроциты. Двигательные нейроциты отличаются непропорционально крупным по сравнению с тонкими разветвленными дендритами телом. Нейроциты ретикулярной формации посылают свои отростки в кору больших полушарий, базальные ядра мозга и мозжечок, объединяя их в общую систему.
Известно, что ретикулярная формация регулирует возбудимость и тонус всех отделов ЦНС. В свою очередь, кора большого мозга контролирует и регулирует его деятельность благодаря большому количеству кортикоретикулярных связей.
Ретикулярная формация отличается высокой чувствительностью к ряду гуморальных раздражителей (углекислота, адреналин) и многим фармакологическим средствам. Она участвует в регуляции многих автономных функций: в поддержании состояния бодрствования, контроле рефлекторной деятельности спинного мозга и т. д.
Область таламуса. Выделяют около 30 ядер:
• передние ядра (3–5), являющиеся подкорковыми центрами обоняния;
• вентромедиальные ядра (10–11), формирующие подкорковые центры общей чувствительности;
• медиальные ядра (2–3), образующие подкорковый центр экстрапирамидной системы. Это интеграционный центр промежуточного мозга;
• срединные ядра (4–5), являющиеся подкорковыми центрами вестибулярных и слуховых функций;
• задние ядра (7–8) – подкорковый центр зрения;
• ретикулярные ядра – чувствительные ядра, связанные с ядрами ретикулярной формации ствола головного мозга.
Гипоталамус. Это скопление более 30 пар ядер. Филогенетически это древнее образование, оно имеется у всех хордовых животных, хорошо развито даже у рыб. Максимума развития и дифференцировки гисталамус достигает у млекопитающих: у них особенно развито серое вещество, формирующее ядра.
Ядра гипоталамуса связаны не только между собой, но и с другими отделами головного мозга: обонятельной луковицей, аммоновым рогом, таламусом. Они имеют мощную капиллярную сеть – на 1 мм приходится от 1100 до 2600 капилляров (в коре мозга только 440–900). Сосуды, питающие гипоталамус, отличаются высокой проницаемостью стенок. Имеются сосудистые и нервные (гипоталамо‑гипофизарный пучок) связи между ядрами гипоталамуса и нейрогипофизом.
Гипоталамус – это высший центр вегетативных функций организма, который координирует нервную и гуморальную регуляцию деятельности всех внутренних органов;
– осуществляет регуляцию сердечно‑сосудистой деятельности, температуры тела, выделения слюны, соков тонкой кишки, мочи, пота и др.;
– участвует в организации поведения, удовлетворяющего основные биологические потребности организма. Здесь располагаются центры: голода (серый бугор), насыщения, терморегуляции, жажды, водного насыщения, полового и родительского поведения, агрессии и страха (задний гипоталамус);
– организует эмоциональное поведение путем активации центров положительного подкрепления (положительные эмоции) и отрицательного подкрепления (отрицательные эмоции);
– запускает на основе анализа внешней среды три группы реакций:
1) вегетативной нервной системы, направленные на поддержание гомеостаза;
2) поведенческие, направленные на удовлетворение потребностей;
3) эндокринной системы.
Выяснено, что задние ядра гипоталамуса влияют на деятельность симпатической, а передние – парасимпатической нервной системы. Ядра гипоталамуса, относящиеся к передней группе, выделяют нейросекреты – гормоны (вазопрессин, окситоцин), которые по аксонам нейроцитов этих ядер, проходящих в ножке гипофиза, попадают в его заднюю долю. Средние ядра регулируют различные стороны обмена веществ, активизируя аденогипофиз с помощью специальных рилизинг‑факторов (либерины и статины).
Мозжечок. В мозжечке различают червь и два полушария, разделенные бороздами на дольки. Дольки полушария разделены бороздками на листочки – извилины мозжечка.
Червь мозжечка состоит из белого вещества, в котором проходят центростремительные и центробежные волокна, и участков серого вещества, расположенных в толще белого в виде 4 пар ядер: зубчатого, переднего межпозиционного (пробковидного), заднего межпозиционного (шаровидного) и ядра шатра.
Поверхность мозжечка покрыта серым веществом – корой. В центре каждой извилины находится слой белого вещества, в составе которого проходят волокна, связывающие ядра червя с корой и кору с червем.
В коре мозжечка различают 3 слоя: молекулярный, слой клеток Пуркинье (ганглиозный) и зернистый (рис. 9.2).
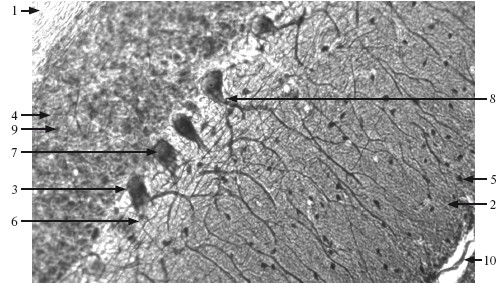
Рис. 9.2. Мозжечок. ×400.
1 – белое вещество; 2 – серое вещество (молекулярный слой коры мозжечка); 3 – слой клеток Пуркинье; 4 – зернистый слой; 5 – звездчатые клетки; 6 – корзинчатые клетки; 7 – клетки Пуркинье; 8 – главный дендрит клетки Пуркинье; 9 – клетки‑зерна; 10 – мягкая мозговая оболочка.
Молекулярный слой состоит из звездчатых и корзинчатых нейроцитов. Звездчатые нейроциты располагаются по всей толще молекулярного слоя. От тела клетки отходят один‑два, редко три дендрита. Аксоны звездчатых нейроцитов образуют синапсы с дендритами клеток ганглиозного слоя.
Корзинчатые нейроциты располагаются в глубоких слоях молекулярного слоя. От тела клетки отходят несколько дендритов и аксон, который тянется параллельно поверхности извилины. По своему ходу аксон отдает восходящие и нисходящие коллатерали. Восходящие коллатерали разветвляются в различных слоях молекулярного слоя, а нисходящие подходят к клеткам ганглиозного слоя, образуя вокруг их тел сплетения корзинчатой формы.
Слой клеток Пуркинье образован телами грушевидных нейроцитов, располагающихся в один ряд. Эти клетки впервые были описаны чешским гистологом Я. Пуркинье. Они имеют грушевидное тело размером до 60 мкм в длину и до 35 мкм в ширину. От их вершины отходит 2–3 дендрита, распадающихся в молекулярном слое на большое количество ветвей. Ветвление их происходит в плоскости, перпендикулярной к длине извилин. От основания клетки отходит аксон, направляющийся через белое вещество в ядра червя. От нейрита отходят коллатерали, вступающие в контакт с телами и дендритами соседних грушевидных нейроцитов.
Зернистый слой состоит из нейроцитов‑зерен, звездчатых и горизонтальных веретенообразных нейроцитов. Нейроциты‑зерна составляют основную массу нейроцитов мозжечка. Это самые маленькие нервные клетки. Их диаметр 5–6 мкм. От тела клетки отходят 3–4 дендрита. Аксон направляется в молекулярный слой, в котором Т‑образно делится на 2 ветви, идущие параллельно поверхности извилины. Аксоны нейроцитов‑зерен вступают в контакт с дендритами грушевидных, звездчатых и корзинчатых нейроцитов.
Различают звездчатые нейроциты с длинными и короткими аксонами. Короткие аксоны звездчатых нейроцитов располагаются в поверхностных слоях зернистого слоя. От их тела отходят сильно ветвящиеся дендриты, часть которых направляется в молекулярный слой и контактирует с аксонами клеток‑зерен. Аксоны ветвятся в зернистом слое и вступают в синаптические связи с дендритами различных нейроцитов этого слоя.
Длинные аксоны звездчатых нейроцитов имеют дендриты, обильно ветвящиеся в зернистом слое. Аксоны входят в белое вещество, обеспечивая связи с другими областями коры мозжечка.
Веретенообразные горизонтальные нейроциты располагаются под слоем грушевидных нейронов. От тела клетки отходят дендриты, идущие в горизонтальном направлении и разветвляющиеся в зернистом и ганглиозном слоях. Аксоны клеток направляются в белое вещество.
Нейроциты молекулярного и зернистого слоев являются ассоциативными. Они участвуют в образовании сложных сочетательных систем коры мозжечка.
Информация в кору мозжечка поступает по так называемым лазающим (восходящим), моховидным и параллельным нервным волокнам. Лазающие волокна из белого вещества направляются к телам грушевидных нейронов, участвуя в образовании корзинок, и наподобие лиан обвиваются вокруг их дендритов.
Моховидные волокна многократно разветвляются в зернистом слое, вследствие чего их концевые ветвления напоминают мох. Эти волокна вступают в контакт с дендритами нейроцитов‑зерен. Места контактов имеют вид неправильной формы телец и получили название клубочков мозжечка.
Грушевидные нейроны получают афферентные импульсы или непосредственно через восходящие волокна, или посредством нейроцитов‑зерен, дендриты которых контактируют с моховидными волокнами, а аксоны в молекулярном слое – с дендритами грушевидных клеток, или через аксоны корзинчатых и звездчатых нейроцитов, дендриты которых вступают в контакт с аксонами клеток‑зерен. Грушевидные нейроциты являются единственными клетками коры мозжечка, аксоны которых выходят за пределы коры, осуществляя синаптические связи с ядрами червя.
Нейроглия коры мозжечка представлена макрофагической глией, астроцитами, олигодендроцитами и особыми глиальными клетками, характерными только для коры мозжечка, так называемые глиоцитами с султаном. Эти клетки имеют опорное значение, расположены в зернистом слое, а их многочисленные отростки пронизывают молекулярный слой, поддерживая дендриты грушевидных клеток.
Дата добавления: 2019-02-12; просмотров: 650; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!