Эдвардс синдромы— трисомия 18. 4 страница
10. 12. Митоздың негізгі кезендері.
Митоз - соматикалық жасушалардың бөлінуі. Осы әдіс генетикалық материалдың жас жасушаларға тең бөлінуін және жасуша ұрпақтарындағы хромосоманың ұқсастығын қамтамасыз етеді.
Митоздың биологиялық маңызы - хромосома санының екі еселенуі және олардың жас еншілес жасушаларға тең бөлінуі. Митоз процесі 4 сатыдан өтеді: профаза, метафаза, анафаза және телофаза.
· Интерфаза – жасушаның екіге бөлінуі аралығындағы дайындық кезеңі. Бұл кезеңде боялып бекітілген ядроның боялған жіңішке жіпшелерден тұратын торлы құрылымын байқауға болады.Интерфаза G1,S,G2 бөлім, кезеңдерінен тұрады.
· G1- кезеңінде жасуша ішінде метаболизм үдерісі қарқында жүзеге асады.
S- кезеңінде жасушадағы генетикалық кодты сақтайтын ДНҚ репрликациясы болады яғни ДНҚ екі еселенеді. G2- кезеңінде жасушадағы органойдтар екі еселенеді. Сонымен интерфаза кезеңінде жасуша митозға толығымен дайындалады. Профаза – ядроның бөлінуге дайындалған бірінші сатысы. Бұл кезде хромосома жіпшелерінің өз осінде шиыршықталып бұратылуының салдарынан хромосомалар қысқарып, жуандайды. Профаза кезінде хромосомалар кариолимфада кездейсоқ жерлерде орналасады, осы кезеңде ядрошықтар бұзылады. Ал профазаның аяқ кезінде ядро қабығы бұзылады да хромосомалар цитоплазма мен кариоплазманың сұйық заттарының қосындысы – миксоплазманың ортасында қалады.
|
|
|
Метафаза фаза сатысының екі кезеңі бар: метакинез – хромосомалар жасушаның экватор аймағына жиналып шоғырланады, жасуша бөлінуге дайындалады; нағыз метафаза – хромосома жіпшелері центромералармен байланысады, хромосомалар хроматидтерге жіктеледі. Клетка цитоплазмасы бұл кезде тұтқырлығын жоғалтады. Бұл кезеңде әрбір хромосоманың центромерасы дәл экваторда, ал қалған денесі экватордан тыс жазықта болуы мүмкін.
Анафаза – хроматин жіпшелерінің болашақ жас жасушалардың полюстеріне қарай созылып, ахроматин ұршығын құрау кезеңі. Ахроматин ұршығының белдеуінде хромосомалар түрліше орналасып, ең алдымен аналық жұлдызын құрайды. Сонан кейін аналық жұлдыздағы қосарланып орналасқан гомологиялық хромосомалардың ұзынынан бөлінуі нәтижесінде пайда болған жас хромосомалар жасушаның полюстеріне қарай ығысады да, екі жас хромосома жұлдыздары пайда болады. Осының нәтижесінде бір жасуша екіге бөлініп, жас жасушалар түзіледі. Анафазада хромосомалардың жасуша полюсіне ығысуы бірден басталып, өте тез арада бітеді.
|
|
|
Телофаза митоздың ақырғы сатысы. Телофаза кезінде хромосомалардың қозғалуы аяқалады, митоздық аппарат бұзылады, ядрошықтар пайда болады. Клетканың қарама-қарсы полюсінде жаңа пайда болған хромосомалардың сыртынан ядролық қабық пайда болады. Жаңа ядролардың қайта құрылуымен қатар әдетте клетка денесі бөлініп, цитотомия не цитокинез өтеді де, екі жасуша құрылады. Телофаза кейде цитотомиямен аяқталмай екі ядролы жасуша құрылады.
11. Клетка айналымының фазалары
Жасушаның тіршілік циклі немесе Митоздық айналым деп жасушаның пайда болуы, құрылысының күрделенуі, атқаратын қызметіне икемделуі, бөлінуі немесе тіршілігін жою кезеңдері аралығындағы әрқилы құбылыстар жиынтығын айтамыз. Яғни, жасуша циклі дегеніміз жасушаның митоздық екі бөлінуі арасындағы байқалатын құбылыстар жиынтығы.
Ол 4 кезеңге бөлінеді:
|
|
|
1. пресинтетикалық
2. синтетикалық
3. постсинтетикалық
4. митоз
Пресинтетикалық кезеңде жаңадан пайда болған жас жасуша өсіп аналық жасушаның мөлшеріне жетеді. Ол үшін жасушада құрылыс материалының(көмірсу,май және белок) синтезделуі қажет.
Синтетикалық кезеңде ДНҚ молекуласы синтезделінеді де тұқым қуалаушылық материал 2п 4с күйінде кездеседі.
Постсинтетикалық кезеңде жасуша бөлінуге дайындалады. Ол үшін жасушада көмірсу, май және белоктар синтезделіп, олардың қоры көбее түседі, органеллалар саны да көбееді,себебі келесі кезеңде (митоз) олардың бәрі де екіге бөлінуі қажет.
Митоз - жасушаның бұрыс бөлінуі. Митоз дене жасушаларында байқалады. Митоз процессінде ядро күрделі өзгерістерге ұшырайды, жаңадан пайда болған жасушаларға тұқым қуалаушылық материалы тепе-тең бөлініп беріліп отырады. Бұл митоздың биологиялық маңызы болып табылады.
Митоз 4 фазадан тұрады: профаза, метафаза, анафаза, телофаза.
1. Профазада ядро көлемі ұлғайып, хроматин жіпшелері тығыз ширатылып, жуандап, қысқарып митоздық хромосомаларға айналады. Профаза аяғында бөлінуші жасушаның екі полюсінде жасуша орталығы пайда болып, ахроматин жіпшелерімен жалғасып тұрады.
|
|
|
2. Метофазада хромосомалар өздерінің центромералары арқылы ахроматин жіпшелеріне жалғанып бөлінуші клетканың ортан беліне- экваторына шоғырланады. Бұл кезеңде әрбір хромосома екі хроматидаға ажырап, тек центромера арқылы байланысып тұрады. Олардың пішіні Х таңбасына ұқсас болады.
3. Анафазада хромосомалардың хроматидалары бір-бірінен толық ажырасады да әртүрлі полюстерге қарай тартыла бастайды.
4. Телофазада бөлінуші жасушаның полюстеріне жиналған хромосомалар жіңішкеріп, ұзарып хроматин жіпшелеріне айналады, ядро шырыны, ядрошық, ядро қабықшасы пайда болып екі жаңа ядро түзіледі. Ядро екіге бөлінгеннен кейін цитоплазмада екіге бөлінеді.Сөйтіп, митоз негізінде бір жасушадан жаңа екі жасуша пайда болады.
13. Амитоз
Клетканың бөлінуі – барлық тірі ағзалардың көбеюлері мен жеке дамуы негізінде жатқан биологиялық үрдіс. Клетканың көбеюі әр уақытта ядроның бөлінуінен басталады. Мұның мынадай үш түрі бар: амитоз, митоз және мейоз.
Амитоз (amitosis; гр. а — жоқ; гр. mitos — жіп) - бөліну кезінде жасуша ядросында күрделі өзгерістер жүрмейтін жасуша бөлінуінің сирек кездесетін түрі. Амитоздық бөліну кезінде жасуша хромосомалары көрінбейді және бөліну ұршығы түзілмейді. Алдымен ядро, соңынан цитоплазма бөлініп, бір аналық клеткадан бірнеше жас клеткалар пайда болады. Адам мен жануарлар организмдерінде амитоздың үш: генеративті, дегенеративті және реактивті түрлері кездеседі. Генеративті амитоз нәтижесінде пайда болған жас жасушалар, қалыпты жағдайда, бөлінуден кейін одан әрі қарай митоз арқылы белініп көбейе береді. Реактивті амитоз — адам мен жануарлар организмдеріне кездейсоқ факторлардың эсер етуі нәтижесінде, мысалы, жарақаттану ошақтарында байқалады. Жарақаттанған ұлпа жасушалары жедел реактивті амитоз арқылы бөлініп, жазылғаннан кейін, ұлпа жасушалары одан әрі митоз арқылы бөліне береді. Дегенеративті амитоз ұлпа жасушаларында тіршілік тоқтап, оларда ыдырау процесі басталған кезінде байқалады.
14-15. Эндомитоз және политения
Эндомитоз. Эндомитоз деп клеткадағы хромосоманың репродукциялануы кезінде ядроның бөлінбей жүру процесін айтады. Осының нәтижерінде клеткада хромосома саны көбейеді, кейде ол бастапқы санымен салыстырғанда ондаған есе артады. Эндомитоз өсімдіктерде сияқты жануарлардың да әр түрлі тканьдердегі белсенді жұмыс істейтін клеткаларында кездеседі. Кейде хромосомалардың пайда болуы, клетка санының өсуінсіз өтеді. Бұл кезде әр хромосома еселеп өседі, бірақ жас хромосома өзара байланысқан күйінде қалады. Бұл құбылыс политения деп аталады. Ол эндомитоздың жеке жағдайынан тұрады. Политенді хромосомада жіпшелер саны 1000—2000 шамасына жетеді. Бұл кезде аса зор алып хромосомалар түзіледі. Политения құбылысы бірқатар диффереицияланған тканьлеткаларда байқалады және ол клетка ядросының ерекше байланысты болады.
16. Мейоздың кезендері

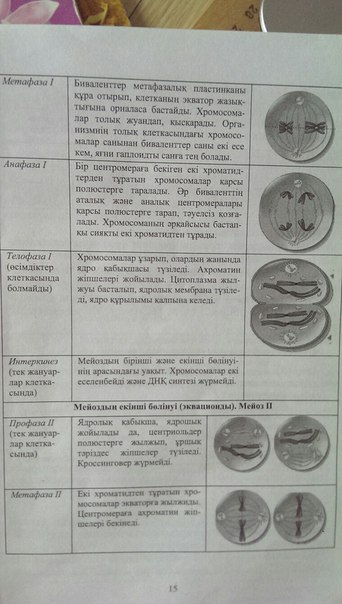

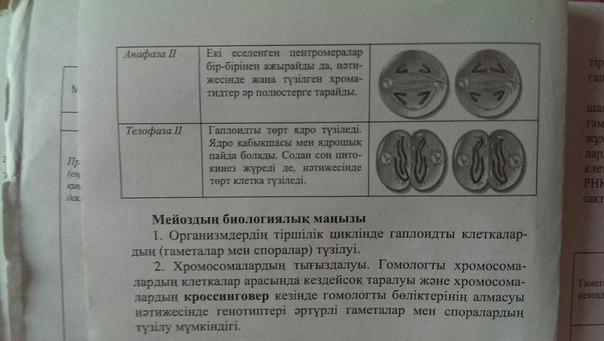
17. Микротүтікшелер, құрылымы, функциялар.
Цитоқаңқа немесе клетка қаңқасы үш компоненттен түзілген. Оларға микротүтікшелер, микрофиламенттер және аралық филаменттер жатады. Микротүктер – клетканың беттік кешенінің туындылары. Олар клетка бетін ұлғайтып, сыртқы ортадан заттарды сіңіру қызметін атқарады.
Микротүтікшелер клетканың бүкіл цитоплазмасын тесіп өтеді. Әрбір микротүтікше диаметрі 20-30 нм іші қуыс цилиндр тәрізді. Олардың қабырғалары 6-8 нм. Микротүтікшелер қабырғасы спираль түрінде бірінің үстінде бірі бұралаңдаған 13 жіпше – протофиламенттерден құралған. Осындай әрбір жіпше тубулин ақуыздарының димерлерінен құралады. Әрбір димер a және b – тубулиннен түзілген. Тубулиндер синтезі түйіршікті эндоплазмалық тордың мембраналарында жүреді, ал спираль клетка орталығында жасалады.Көптеген микротүтікшелер центриольдерге қатысты радиальді бағытта болады. Осыдан барып олар цитоплазмаға тарайды. Олардың бір бөлігі плазмалемманың астында орналасады да, сол жерде микрофиламенттердің шоқтарымен бірге терминальды тор құруға қатысады.Микротүтікшелер өте мықты, олар цитоқаңқаның сүйеуші құрылымын құрайды. Микротүтікшелердің бір бөлігі жасушаға әсер етуші қысым және тарту күштеріне байланысты орналасады.Микротүтікшелер клеткаішілік зат тасымалдауға қатысады. Микротүтікшелердің қабырғасына өздерінің бір ұшымен қысқа тізбек түзетін ақуыз молекулалары жалғасады, олар белгілі бір жағдайларда өздерінің кеңістіктегі конфигурацияларын өзгерте алады. Бейтарап жағдайда тізбек қабырға бетіне параллель орналасады. Бұл жағдайларда тізбектің бос ұшы гликокаликстегі бөлшектермен қосылыс түзулері мүмкін.Осындай бөлшектерді қосқаннан кейін ақуыз өз конфигурациясын өзгертіп, қабырғадан алшақтайды да өзімен бірге бөлшектерді ала кетеді, алшақтаған тізбек бөлшекті өзінің үстінде жатқан тізбекке, ол өзінен кейінге т.с.с. тасымалдайды.Микротүтікшелерде конфигурацияланатын сыртқы тізбектер бар болғандықтан, олар клеткаішілік белсенді тасымалдаудың негізгі ағыстарын түзеді.Микротүтікшелердің қабырғаларының құрылымдары әр түрлі әсер етулерден өзгерулері мүмкін. Бұл жағдайларда клеткаішілік тасымалдау бүлінеді.
Қызметі.Биологиялық мембраналар клетканы тек қана сыртқа ортадан бөліп қоймай, сондай-ақ, өздерінің құрылымдары мен қызметтік ерекшеліктері бар, жеке-жеке аймақтарға – компартменттерге бөледі. Осындай компартменттермен байланысқан мембраналардың өздеріне сай ерекшеліктері болады. 7 мкм-ге жететін аса ұзын микротүктерді стереоцилиялар деп атайды. Олар кейбір, мысалы, есту және тепе-теңдік мүшелерінің сенсорлы клеткалары сияқты арнайы клеткаларда кездеседі. Олардың бастапқы сору қызметтері жойылады да, қызметтері алғашқы орналасуларын өзгертулерімен байланысты болады. Клетка бетінің конфигурациясының өзгеруі клетканы қоздырады, қозу нерв талшықтарының ұштарына беріледі, ал сигналдар орталық нерв жүйелеріне барады. Стереоцилияларды микротүктердің модификациясынан туындаған арнайы органеллалар ретінде қарастыруға болады
18. Когезинтер. Синаптонемді кешен.
Когезин — это комплекс белков, который регулирует процесс разделения сестринских хроматид в ходе деления клетки (как мейоза, так и митоза).
Структура
Когезин состоит из четырех составляющих: Scc1, Scc3, Smc1, Smc3. Белки Smc имеют две главные характеристики: АТФазную активность домена, образующегося при взаимодействии С-конца и N-конца белка) и петлеобразную зону, позволяющую димеризацию Smc. Домен АТФазы и петля соединяются между собой через длинную антипаралелльную "спираль из спиралей". Общая структура димера имеет центральную петлю, окруженную АТФазами. В присутствии АТФ два домена АТФазы могут связываться, формируя структуру кольца. Гидролиз АТФ может вызвать открывание или закрывание кольца.
Scc1 и Scc3 соединяют АТФазные домены Smc1 и Smc3, стабилизируя структуру кольца. N- и C-концы Scc1 соединяются с Smc1 и Smc3. Когда Scc1 соединяется с белком SMC, Scc3 также может присоединиться, соединяясь с С-концевым участком Scc1. Когда Scc1 соединяется и с Smc1, и с Smc3, комплекс когезина формирует закрытую форму кольца. Если же он соединяется только с одним из белков SMC, то образуется структура открытого кольца. Недавно было обнаружено, что кольца когезина могут димеризироваться, образуя структуру из двух колец, скрепленных частью Scc3 в форме наручников, причем в каждом кольце проходит нить ДНК.
Функции. У кольца когезина есть три функции:
Используется для удержания сестринских хроматид, соединенными друг с другом в течение метафазы, гарантируя их соединение в процессе митоза(мейоза). После разделения две хроматиды движутся к противоположным полюсам клетки. Без когезина клетка не смогла бы контролировать изоляцию сестринских хроматид.
Облегчает присоединение веретена деления к хромосомам
облегчает восстановление ДНК путем рекомбинации
Синаптондық кешені (СК) - мейоз кезінде гомологиялық хромосомалардың (апалы-сіңлілі (сестринских) екі жұп хромосомалар) арасындағы қалыптасқан ақуыз кешені.
СК хромосомалардың шағылысу кезінде арасындағы байланыстырушы буын ретінде әрекет етеді
СК 1956 жылы Montrose J. Moses өзен шаянының алғашқы сперматазоидтарынан, сондай –ақ D. Fawcett көгершін,мысық пен адамның сперматозоидтарынан сипаттады.
СК генетикалық рекомбинацияға қажетті емес екендігі қазір уақытта дәлелденген. Зерттеу барысында оның рекомбинациядан кейін қалыптасатыны, ал СК-ны құруға қатыспайтын ашытқының мутантты жасушасы генетикалық материал алмасуына қатысуын жалғастырады. (яғни рекомбинация). Қазіргі уақытта СК гомологты хромосомалар арасындағы қалыпты кроссинговерге «тірек» ролін атқарады.\
19. Кариотип. Кариотиптік.
Кариотип – берілген биологиялық түр мен берілген ағзаның клеткаларына немесе клетка-ың линиясына тән толық хромосомалар жиынтығының белгілер жиыны (мөлшері,көлемі,пішіні т.б.)
Кариотип термині 1924ж. Г.А.Левитскиймен енгізілді. Кариотип - бір түрдің жасушасына тән толық хромосомалық жинағының морфологиялық ерекшеліктерінің жиынтығы.
Кариотиптің түрарнайылығы хромосоманың жалпы санымен анықталады, олардың морфологиялық ерекшеліктерімен, сонымен қатар дифференциалды бояу әдісі арқылы анықталатын олардың функционалды құрылымдық ұйымдасуымен анықталынады.
20. Адам кариотипінің ерекшеліктері.
Кариотипті сипаттау үшін барлық хромосомалық жинақтың гомологиялық хромосомасының видеосуреті пайдаланылады, яғни кариограмма, немесе әр гомологтың кескіндік суреті — идеограмма қолданылады. Хромосомалық анализ нәтижесі болып кариотипті формула түрінде хромосома санын көрсету арқылы, жыныс хромосомасының жинағын және хромосомалық аберрацияны көрсету арқылы сипаттау табылады (егер олар анықталса).
Адамның кариотипі қалыпты жағдайда 46 хромосомадан тұрады. Бұндай диплойдтық сан ата-аналық гаметалардың бірігуінің нәтижесінде туындайды, ал ата-аналық гаметалардың әр қайсысы 23ке тең хромосоманың гаплойдты санынан тұрады. Осылай адам ұрығы зигота сатысынан 23 жұп хромосомаға ие болады, олардың 22-сі аутосомалы және бір жұбы- жыныс хромосомадан немесе гоносомадан тұрады (Х және Y). Екі жыныс Х хромосомасының болуы (кариотип 46,ХХ)аналық жыныс хромосомасынасай, Х- және Y-хромосоманың болуы (кариотип 46,ХY) —аталыққа сай келеді.
Центромерлік аудандар (С-сегменттер) адам кариотипіндегі кез келген хромосоманың міндетті құрылымдық компоненті болып табылады. Алайда С-блоктардың көлемі бойынша анық жеке аралық полиморфизм байқалады, ол прицентромерлік аудандардың әртүрлі өлшемдерімен шартталған С-сегменттердің құрамына сателитті ДНҚ-ның жай және күрделі нуклеотидтік қайталамалары кіреді.Адамның хромосомасының әр жұбында соның өзіне ғана тән альфоидты ДНК-сы болады.
Адам кариотипіндегі акроцентрлік аутосоманың бес жұбының қысқа иықтарының құрылымдық компоненті болып спутниктер(s) және спутниктік жіптер табылады (stk) (англ. satellites және satellite stalks сәйкесінше). Полиморфты нұсқалар арасында сонымен бірге қос спутникті және қос ядрошық түзуші аудандардан тұратындары да кездеседі (ЯТА). Спутниктер сатДНҚ ретінде көрсетілген. Спутниктік жіптер N- немесе Ag-NOR блоктармен белгіленеді.ЯТА-да көпрет қайталанатын рибосомалық гендердің кластерлері орналасқан. Әр акроцентрлік хромосома р-генннің әртүрлі санын құрайды.
21. Хромосомалардың центромерлік және теломерлік аймақтары.
Хромосомалар негізінен ДНҚ және ақуыздардан нуклеопротеинді кешен – хроматин түзеді. Хроматиндерді, олай атау себебі негізгі бояғыштармен боялу қасиетіне байланысты. Хроматиннің митотикалық суперкомпактизациясы жарық микроскопы арқылы хромосомалардың сыртқы түрін көруге мүмкіндік береді. Митоздың бірінші жартысында олар хромосоманың ерекше ұйымдасқан бөлігі біріншілік перетяжка(центромера немесе кинетохор) ауданында өзара байланысқан екі хроматидтен тұрады. Митоздың екінші жартысында хроматидтердің бір-бірінен ажырауы жүреді. Олардан жаңа жасушалармен бөлінетін біржіпшелі жаңа хромосомалар пайда болады.
Центромераның орналасуына және екі жағында орналасқан иықтарының ұзындықтарына қарай хромосоманың бірнеше формаларын бөледі: теңиықты,немесе метацентрлі (ортасында центромерасы бар), теңиықты емес, немесе субметацентрлі (центромерасы екі ұшының біреуіне қарай жылжыған, таяқтәріздес, немесе акроцентрлі (центромерасы хромосоманың ұшына қарай ораналасқан, және нүктелік —өте кішкентай, формасын анықтау өте қиын.Рутинді әдістермен хромосомаларды бояғанда олар формалары бойынша және мөлшерімен ерекшеленеді.
Дифференциалды бояу әдістерін қолданғанда бірдей емес флуоресценция немесе әр хромосома мен оның гомологына қатаң спецификалы хромосоманың ұзындығын бойлай бояудың таралуы анықталады. Біріншілік перетяжка (центромера) ауданында формасы диск тәріздес пластинді құрылым – кинетохор орналасқан. Оған центроильдерге бағытталған митотикалық ұршықтың микротүтікшелер шоғыры келеді. Осы микротүтікшелер шоғырлары митоз кезінде жасуша полюсына хромосомалардың жылжуына қатысады.Әдетте әрбір хромосоманың бір ғана центромерасы болады(моноцентрлі хромосомалар), бірақ дицентрлі және полиценрлі хромосомалар кездесуі мүмкін,яғни көптеген кинетохорасы бар.Центромера маңында маңызды, центромералы, сатиллетті ДНҚ бар, ол нуклеотидті тізбектердің қайталануының жоғарылығымен.Кейбір хромосомалар екіншілік перетяжкасы болады. Кейінгісі әдетте хромосоманың ұшына қарайц орналасқан және кіші ауданды алады, спутник. Екіншілік перетяжкаларды, сонымен қатар ядрошықтық ұйымдастырушылар деп те атайды, себебі хромосоманың осы бөліктерінде интерфазада ядрошықтың түзілуі болады. Осы жерде рРНҚ-ң синтезіне жауапты ДНҚ орналасқан. Адам хромосомаларында ядрошықтық ұйымдастырушылар центромераларға жақын қысқа иықтарында орналасқан. Хромосомалардың иықтары соңғы бөлігі – теломералармен аяқталады. Хромосомалардың теломерлік ұштары теломерлік бөліктерінен айырылған (үзілулер нәтижесінде) хромосома ұштарымен салыстырғанда басқа хромосомалармен немесе фрагменттерімен қосылу қасиетине ие емес. Теломераларда ДНҚ синтезі процесі кезінде хромосоманы қысқарудан сақтайтын ерекше теломерлі ДНҚ локализденген.
22. Адам хромосомасының сегменттерінің молекулалық гетерогенділігінің қысқаша сипаты.
Дата добавления: 2016-01-04; просмотров: 18; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!