Смена места жительства и ископаемые гены опсинов
На волне огромного интереса к латимерии к местам ее естественного обитания были направлены экспедиции. Спускаясь на глубину, ученые наблюдали этих рыб в подводных пещерах вблизи Коморских островов и у берегов Южной Африки. Днем рыбы прячутся в пещерах, а ночью медленно перемещаются вблизи дна в поисках пищи. На глубину 100 м и более, где обитают латимерии, проникает лишь очень слабый синий свет.
Необычный образ жизни латимерий и их уникальный статус привлекли внимание к строению их зрительной системы и генов опсинов. Оказалось, что латимерия имеет родопсин, настроенный на восприятие тусклого рассеянного света, но не имеет MWS/LWS‑опсина, регистрирующего свет в красно‑зеленой области, как у других рыб и у человека. Поскольку рыбы, млекопитающие и большинство других позвоночных животных имеют хотя бы одну версию такого опсина, ясно, что предки латимерии также имели этот ген, но на каком‑то этапе эволюции он был утрачен. Потеря этого гена вызывает общий вопрос: как и почему столь важный для одних видов ген теряется у других? Проиллюстрировать картину утраты функций гена можно на примере другого гена опсина, который все еще присутствует в ДНК латимерии, но постепенно разрушается.
У латимерии есть один ген опсина SWS. Вспомним, что этот опсин, настроенный на коротковолновую область спектра, у человека и птиц используется для восприятия фиолетового света, а у некоторых животных – для восприятия ультрафиолетового света. Однако в последовательности ДНК коротковолнового опсина латимерии обнаружено множество замен, которые нарушают информационное содержание гена. Например, в позициях 200–202 этого гена у мышей и других животных расположен триплет CGA, а у латимерии в этом месте стоит триплет TGA. Замена C на T может показаться не очень существенной, но в данном случае это важно. Дело в том, что триплет TGA представляет собой так называемый стоп‑кодон, который служит в качестве точки, прерывающей трансляцию последовательности гена опсина SWS. Это лишает латимерию возможности синтезировать работоспособный белок SWS‑опсин. Кроме того, в последовательности гена есть и другие делеции и замены, которые серьезно нарушают текст. Подобная ситуация говорит о том, что данный ген потерял свою функцию, то есть стал ископаемым геном (биологи называют такие гены псевдогенами, но я буду по‑прежнему называть их ископаемыми). Этот ген функционировал у предков латимерии, но сегодня уже не работает. Его все еще можно узнать по сохранившимся фрагментам, но, поскольку функция его утрачена, он продолжает накапливать мутации и делеции и когда‑нибудь совсем исчезнет из последовательности ДНК латимерии, как исчез ген опсина MWS/LWS (а также один из глобиновых генов у предков ледяной рыбы, о которой мы говорили в первой главе).
|
|
|
|
|
|
Вы, вероятно, спросите: почему хорошему гену было позволено разрушиться? Являются ли ископаемые гены редкой особенностью таких таинственных существ, как латимерия и ледяная рыба? Прежде чем ответить на эти вопросы, я приведу еще один показательный пример.
Исследование генов коротковолновых опсинов у дельфинов и китов показало, что у них, как у латимерии, ген опсина SWS тоже стал ископаемым. Например, у афалины в начале гена опсина SWS не хватает нескольких оснований (рис. 5.1).

Рис. 5.1. Ископаемый ген опсина у дельфина. Показаны короткие фрагменты генов коротковолновых опсинов дельфина и коровы. Закрашенные клетки обозначают позиции, в которых были потеряны основания (звездочки), что привело к нарушению кода опсина у дельфина. Рисунок Джейми Кэрролл.
Это нарушает триплетную структуру последовательности, сдвигает рамку считывания и делает ген полностью нефункциональным. Анализ генов коротковолновых опсинов других китообразных выявил в них ряд других замен, которые также привели к нарушению функций этих генов. У всех дельфинов и китов есть ископаемый ген опсина SWS.
|
|
|
Таким образом, мы имеем два примера ископаемых генов опсина SWS – у латимерии и китообразных. Есть ли между этими животными что‑нибудь общее, что могло бы объяснить потерю функции их коротковолновых опсинов?
Прежде всего нужно сказать, что отмирание генов опсинов у этих групп животных произошло независимо. Мы знаем это, поскольку знаем положение этих видов животных на эволюционном древе позвоночных (рис. 5.2).

Рис. 5.2. Один и тот же ген опсина дважды стал ископаемым. Распределение различных мутаций в генах опсинов SWS латимерии и китообразных, а также эволюционная связь этих видов животных говорят о том, что ген SWS‑опсина становился ископаемым как минимум два раза (звездочки). Рисунок Джейми Кэрролл.
Латимерия относится к группе примитивных рыб, отделившейся от ветви, давшей начало четвероногим позвоночным. У амфибий, рептилий, птиц и многих млекопитающих гены коротковолновых опсинов не пострадали, следовательно, отмирание этого гена у латимерии произошло в ходе эволюции ее ветви. Поскольку гиппопотамы и коровы – близкие родственники китов и дельфинов – сохранили функциональные SWS‑опсины, а китообразные нет, можно сделать вывод, что функция SWS‑опсина китообразных была утеряна у общего предка всех дельфинов и китов. Отсутствие функционального коротковолнового опсина у современных дельфинов и китов связано с наследованием этого ископаемого гена от их общего предка, жившего более 40 млн лет назад.
|
|
|
Лучшим объяснением потери функции отдельных генов является изменение образа жизни животных. Безусловно, должна быть какая‑то связь между неиспользованием опсина SWS и жизнью в море. Дельфины и киты живут только в воде и принадлежат к единственному отряду млекопитающих, полностью потерявших способность различать цвета (поскольку в колбочках их сетчатки есть только один тип опсина, тогда как у большинства других млекопитающих их два). Как мы уже говорили, родопсин дельфинов настроился на восприятие тусклого света в синей области видимого спектра. Латимерия тоже обитает на большой глубине и, по‑видимому, не нуждается в цветовом зрении. Экологическое объяснение потери функции коротковолнового опсина состоит в том, что он перестал быть необходимым предкам этих животных.
Отсутствие потребности в белке привело к тем изменениям, которые мы видим в последовательности его гена. Если опсин больше не нужен, естественный отбор, который в норме должен поддерживать его существование, ослабевает. А если ослабевает давление со стороны естественного отбора, ДНК перестает очищаться от мутаций, разрушающих ее функцию. Случайность возникновения мутаций означает, что все гены подвергаются мутациям. В большинстве случаев вредные мутации исчезают благодаря очистительному действию естественного отбора, поскольку мутантные особи и их потомство являются менее приспособленными. Но если какой‑то признак перестает быть предметом отбора (например, при перемене места жительства), необходимые раньше гены становятся ненужными, и в них накапливаются мутации.
Используй, или потеряешь.
Говоря научным языком, ископаемые гены – это то, во что должны были бы превратиться со временем любые гены в результате постоянного действия мутаций, если бы не было естественного отбора. Очевидным признаком того, что ген стал ненужным, является накопление мутаций, разрушающих содержащуюся в нем информацию. Таким образом, ископаемые гены отражают различия в образе жизни современного вида и его предков, и если такие гены удается обнаружить, они помогают реконструировать ход естественной истории.
Я приведу еще несколько примеров отмирания генов опсинов, связанных с другими переменами в образе жизни животных, а потом мы вовлечем в наше обсуждение еще больше ископаемых генов и еще больше видов.
Жизнь в темноте
Основная теория, объясняющая уменьшение числа генов опсинов и потерю полноценного цветового зрения у предков млекопитающих, заключается в том, что первые млекопитающие, напоминающие грызунов, были ночными животными, и поэтому способность различать цвета была им просто не нужна. У млекопитающих переход к ночному образу жизни происходил многократно, поэтому один из способов проверки этой теории и более общей идеи о том, что потеря функций генов связана со сменой образа жизни, состоит в изучении более современных видов с заметно различающимся образом жизни.
Примером может служить ночная (совиная) обезьяна – единственный представитель высших приматов, ведущий ночной образ жизни (рис. 5.3).
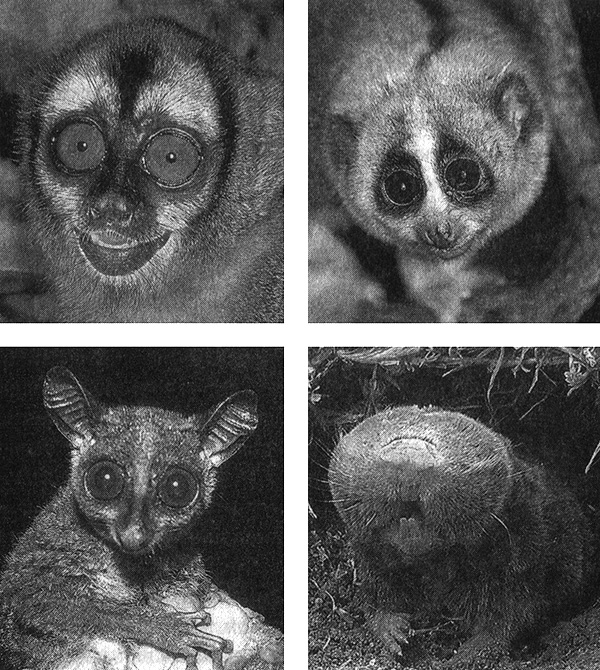
Рис. 5.3. Ночные или подземные млекопитающие с ископаемыми генами опсинов. У ночной обезьяны (вверху слева; фотография Грега и Мэри Бет Димиджан), галаго (внизу слева; фотография Б. Смита, Нигерия), толстого лори (вверху справа; фотография Ларри П. Такетта, www.tackettproductions.com) и слепыша (внизу справа; фотография Тали Кимчи) гены коротковолновых опсинов превратились в ископаемые гены в результате адаптации к ночному или подземному образу жизни.
Анализ генов коротковолновых опсинов этих животных показал, что в них также накопились мутации, из‑за которых они перестали работать. На расстоянии 60 оснований от начала гена опсина триплет TGG мутировал в TGA – еще один случай появления стоп‑кодона, прерывающего трансляцию оставшейся части гена. У всех дневных родственников ночных обезьян ген SWS‑опсина полностью функционален, и это хороший довод в пользу того, что переход к ночному образу жизни привел к отмене давления со стороны естественного отбора.
Гипотезу о связи между ночным образом жизни и опсином SWS можно дополнительно проверить, исследуя гены опсинов ночных полуобезьян. Полуобезьяны – это группа примитивных приматов, к которым относятся лемуры, долгопяты, галаго и лори. Среди лемуров встречаются виды, ведущие как дневной, так и ночной образ жизни, однако толстый лори и галаго – исключительно ночные животные (см. рис. 5.3). Анализ их генов опсинов SWS показал, что эти гены стали ископаемыми. У обоих видов в начале гена коротковолнового опсина отсутствует большой фрагмент текста, что делает невозможным синтез соответствующего белка. Поскольку у обоих видов эта делеция имеет один и тот же размер и находится в одном и том же месте, это означает, что потеря функции опсина SWS произошла у общего предка галаго и лори и была унаследована его потомками.
До сих пор все логично. По‑видимому, освещенность среды обитания коррелирует с утратой (или сохранением) генов, ответственных за цветовое зрение. Но давайте проведем еще одну проверку: как насчет животных, перешедших к подземному образу жизни?
Слепыши – это грызуны, у которых глаза атрофировались сильнее, чем у каких‑либо других млекопитающих (см. рис. 5.3). Судя по окаменелостям, эта группа животных произошла от наземного предка, обладавшего нормальными глазами. Эволюция слепыша сопровождалась многочисленными изменениями анатомии и физиологии. Его глаза настолько малы, что не могут различать изображения. Кроме того, даже если бы глаза остались нормальными, видеть это животное все равно было бы не в состоянии, поскольку его глаза полностью скрыты под слоем кожи и меха. Однако слепыш может определять время суток: в его глазах сохранилась сетчатка, способная распознавать свет, что позволяет животному поддерживать суточный биоритм.
Исследования показали, что у слепыша нормально функционируют два гена опсина – смещенный в красную область спектра пигмент MWS/LWS, настроенный на восприятие света подкожным глазом, а также родопсин, воспринимающий тусклый рассеянный свет. Очевидно, несмотря на атрофированное зрение, естественный отбор продолжает поддерживать функцию этих генов, чтобы животное могло жить в обычном суточном ритме. Однако ген опсина SWS у слепышей стал ископаемым и содержит множество мутаций, нарушающих генетический текст.
Итак, я привел пять независимых примеров вырождения генов коротковолновых опсинов – у латимерии, китообразных, ночной обезьяны, толстого лори и галаго, а также у слепыша. В каждом случае нарушение функции гена коррелировало с изменением образа жизни животного. В каждом случае имел место особый характер повреждения гена опсина SWS. Этот факт, а также то, что эти виды животных принадлежат к различным ветвям эволюционного древа, а их близкие родственники имеют нормальные опсины, говорит о том, что исчезновение функции генов коротковолновых опсинов происходило многократно и независимо на разных этапах эволюции. Таким образом, предположение о том, что ослабление действия естественного отбора на ген ведет к разрушению этого гена, находит убедительное подтверждение. Более того, у всех перечисленных видов животных другие типы опсинов сохраняют свое строение и функцию, что означает, что процесс разрушения генов является крайне избирательным.
Частая утрата гена опсина SWS и связь этого явления с изменением образа жизни говорят о том, что эволюционные изменения часто сопровождаются нарушением функций генов. Теперь давайте обратимся к анализу генома человека, найдем в нем ископаемые гены и посмотрим, что они могут рассказать о наших отличиях от предков.
Не чувствуете, чем пахнет?
Мы увидели немало примеров того, как смена образа жизни приводит к приобретению или к потере генов, отвечающих за зрение. Другие системы восприятия, особенно обоняние, также имеют важнейшее значение для выживания и поведения животных. Достаточно один раз прогуляться в парке с собакой, чтобы понять, в какой степени «видение» мира этими животными определяется их острым нюхом.
Многие другие млекопитающие также обладают прекрасным обонянием, при помощи которого они находят себе пропитание, идентифицируют партнера и потомство, распознают опасность. На протяжении длительного времени механизм улавливания и различения запахов оставался для ученых загадкой. В 1991 году Линда Бак и Ричард Эксел обнаружили семейство генов, кодирующих рецепторы пахучих молекул. Гены обонятельных рецепторов образуют самое крупное семейство генов в геноме млекопитающих. Мышь имеет около 1400 таких генов, а весь ее геном состоит из 25 тыс. генов. Оказалось, что специфичность распознавания запахов связана с тем, что каждая обонятельная нервная клетка имеет лишь один тип (в редких случаях несколько) обонятельного рецептора, настроенного на восприятие конкретной группы пахучих молекул. Восприятие каждого запаха зависит от сочетания рецепторов, участвующих в его распознавании. За разгадку механизма обоняния Бак и Эксел в 2004 г. были удостоены Нобелевской премии в области физиологии и медицины.
Человеческие гены, ответственные за обоняние, изучены очень подробно. Как оказалось, в этой области восприятия человеку по сравнению с мышью похвастаться нечем. Около половины генов обонятельных рецепторов человека стали ископаемыми, не способными производить функциональные рецепторы. Самый разительный контраст между человеком и другими млекопитающими наблюдается в группе рецепторов, кодируемых так называемыми V1r ‑генами. У мыши обнаружено около 160 функциональных рецепторов V1r , тогда как в геноме человека из 200 генов V1r лишь пять сохранили свою функцию. Очевидно, что наш ассортимент обонятельных рецепторов заметно сократился.
Столь высокое содержание ископаемых генов обонятельных рецепторов говорит о том, что мы больше не полагаемся на обоняние в той же степени, что наши предки. И тут возникает два вопроса. Во‑первых, почему мы перестали использовать такую значительную часть наших обонятельных рецепторов? И во‑вторых, когда это произошло?
Чтобы найти ответ, стоит проанализировать долю ископаемых генов обонятельных рецепторов у других приматов и млекопитающих. Профессор Иов Гилад и его коллеги из Вейцмановского института в Реховоте и Института эволюционной антропологии Макса Планка в Лейпциге изучили наборы генов обонятельных рецепторов у человекообразных обезьян, обезьян Старого и Нового Света и лемуров и сравнили их с набором мышиных генов. Они обнаружили удивительную корреляцию между долей ископаемых генов обонятельных рецепторов и эволюцией полноценного цветового зрения. У мышей, лемуров и обезьян Нового Света, не имеющих полноценного цветового зрения, ископаемыми являются лишь 18 % генов обонятельных рецепторов. Однако у колобусов и других обезьян Старого Света эта цифра достигает 29 %, а у человекообразных обезьян (орангутана, шимпанзе и гориллы) – 33 %. Наконец, у человека ископаемыми стали 50 % генов обонятельных рецепторов. Таким образом, доля нефункциональных генов обонятельных рецепторов значительно выше у тех видов, которые наделены полноценным цветовым зрением. Это означает, что эволюция трихроматического зрения, которое позволяет этим приматам обнаруживать еду, партнеров и опасность, ослабила их зависимость от способности распознавать запахи. Ослабление отбора генов обонятельных рецепторов у видов, обладающих трихроматическим зрением, привело к разрушению этих генов. Напротив, животные, которые в основном полагаются на обоняние, гораздо лучше сохранили свой набор генов обонятельных рецепторов.
Существуют и другие физические, поведенческие и генетические признаки, свидетельствующие об ослаблении роли обоняния в жизни человека и других приматов. Вомероназальный (или сошниково‑носовой) орган – орган восприятия, имеющий форму сигары и расположенный в передней части носовой камеры, – у большинства наземных позвоночных служит для обнаружения феромонов. Однако у человека и высших приматов он в значительной степени редуцирован. Упомянутые мною рецепторы класса V1r играют важнейшую роль в обнаружении феромонов. Таким образом, при выборе партнера мы меньше ориентируемся на феромоны, чем другие млекопитающие, возможно, по той же самой причине – поскольку наши предки для выбора партнера и других функций стали больше полагаться на зрительные сигналы.
Поскольку вомероназальный орган и рецепторы V1r у человека и других высших приматов настолько редуцированы, можно предположить, что дегенерации подверглись и другие механизмы, задействованные в передаче информации от обонятельных органов. И это действительно так. Еще один ген, играющий важную роль в работе вомероназального органа, TRPC2, кодирует белок, регулирующий транспорт ионов в сенсорных клетках. У мышей белок TRPC2 полностью функционален и участвует в реакциях на феромоны. Однако у человека и других высших приматов с трихроматическим зрением и большим количеством редуцированных обонятельных рецепторов ген TRPC2 содержит множество мутаций, нарушивших его функцию.
Тот факт, что гены с разными функциями в обонятельной системе стали ископаемыми генами, – поразительное, исчерпывающее подтверждение предположений о том, что должно происходить в результате ослабления действия отбора на какой‑либо признак. Когда целый орган или процесс становится ненужным, гены, участвующие в реализации различных этапов этого процесса, перестают быть объектом отбора и постепенно превращаются в ископаемые. Эволюция вомероназального органа и его отдельных элементов показывает, что целые комплексы генов могут выходить из употребления, распадаться и исчезать. Этот процесс можно наблюдать у некоторых видов, причем иногда он становится массовым. Я приведу два примера из других царств в качестве иллюстрации того, как эволюция убирает то, что больше не используется.
Используй, или потеряешь
Дрожжи и другие грибы играют важную роль в жизни людей. Дрожжи мы используем для производства пива, вина и хлеба, а грибы стали для нас первым источником антибиотиков. Пекарские и пивные дрожжи Saccharomyces cerevisiae давно уже стали излюбленным лабораторным организмом, поскольку их очень легко выращивать. Эксперименты с дрожжами позволили узнать очень многое о росте и делении клеток, об использовании генов и о биохимических основах жизни.
Однако на свете существуют не только старые добрые пивные дрожжи, но и множество других видов дрожжей. Под микроскопом большинство из них очень похожи, однако их способности переваривать питательные вещества и расти в различных условиях зачастую заметно различаются. Распад питательных веществ на полезные компоненты осуществляется в результате последовательности реакций, называемых метаболическим путем. Одним из наиболее хорошо изученных метаболических путей является путь расщепления галактозы у пекарских дрожжей. Большинство организмов в качестве источника энергии используют глюкозу. Однако, когда глюкозы нет, они вынуждены расходовать запасные сахара (например, крахмал) или искать альтернативные источники энергии. В частности, пекарские дрожжи могут использовать другой сахар – галактозу, которую они умеют переводить в глюкозу путем серии ферментативных реакций. В этих реакциях (в этой цепи) задействованы четыре фермента, закодированные четырьмя разными генами. Для того чтобы дрожжи синтезировали эти ферменты только при необходимости и в присутствии галактозы, синтез этих четырех ферментов контролируют еще три белка. Таким образом, в метаболизме галактозы в пекарских дрожжах участвует семь генов.
Большинство ближайших родственников пекарских дрожжей также могут использовать галактозу, за исключением одного вида дрожжей. Этот вид, Saccharomyces kudriavzevii (попытайтесь быстро произнести это название несколько раз подряд[14]), был обнаружен в Японии, на гниющей листве, тогда как большинство других дрожжей обитают в богатых сахарами средах. Когда мой аспирант Крис Хиттингер занялся изучением генов метаболизма галактозы у S. kudriavzevii, он быстро понял, почему этот организм не может расщеплять галактозу: все гены были испорчены. Все семь генов имели различные повреждения, нарушавшие целостность заключенной в них информации.
У S. kudriavzevii наблюдается разительный контраст между состоянием семи генов, участвующих в расщеплении галактозы, и состоянием их ближайших соседей по ДНК. Соседние гены прекрасно сохранились и так же функциональны, как у пекарских дрожжей и других родственных видов. Если представить себе последовательность гена в виде абзаца текста, то текст каждого гена метаболизма галактозы у S. kudriavzevii во многих местах оказался стертым, тогда как предыдущие и последующие абзацы прекрасно пропечатаны. Это говорит о том, насколько специфичным является процесс отмирания генов. В генах, которые больше не нужны и не используются, накапливается множество мутаций, а нужные соседние гены остаются в целости и сохранности. Судьба семи генов S. kudriavzevii демонстрирует способность естественного отбора сохранять то, что нужно организму, и его бездействие в отношении того, что уже не нужно. Этот вид дрожжей адаптировался к использованию других источников сахара и утратил способность расщеплять галактозу. Без постоянного надзора со стороны естественного отбора гены метаболизма галактозы собрали множество мутаций, стали ископаемыми и постепенно исчезают.
Утрата семи функционально связанных генов кажется мне чрезвычайно выразительным примером того, как значительное число генов могут погибнуть без внимания со стороны естественного отбора, но бывают примеры и более яркие – например, то, что произошло с некоторыми другими микроорганизмами, в частности с патогенным микроорганизмом Mycobacterium leprae, вызывающим проказу.
Анализ генома M. leprae показал, что в нем содержится около 1600 функциональных генов и почти 1100 ископаемых генов – доля мертвых генов тут намного выше, чем в любом другом организме. Бактерия M. leprae является родственником бактерии M. tuberculosis, вызывающей туберкулез легких. Однако M. tuberculosis имеет около 4 тыс. функциональных генов и лишь шесть ископаемых. Сравнение двух видов показывает, что в ходе эволюции около 2 тыс. генов M. leprae оказались испорченными или утерянными. Чем объясняется такое поразительное различие в числе функциональных и ископаемых генов у двух родственных организмов?
Дело в том, что M. leprae ведет совершенно иной образ жизни, чем ее родственник. Эта бактерия может жить только внутри клеток организма‑хозяина. Она поселяется в клетках, называемых макрофагами, и инфицирует клетки периферической нервной системы, разрушение которых приводит к типичным для проказы физическим увечьям. Эта бактерия растет медленнее, чем любые другие известные бактерии (ее клетки делятся примерно раз в две недели, тогда как клетки E. coli у нас в кишечнике делятся каждые 20 минут). Несмотря на многолетние попытки ученых, в лабораторных условиях эту бактерию так и не удалось вырастить в самостоятельном виде. Жизнь внутри чужих клеток позволяет M. leprae пользоваться многими метаболическими процессами хозяина. Поскольку большую часть работы выполняют гены хозяина, многие гены M. leprae оказались без присмотра со стороны естественного отбора. Такое же массовое разрушение генов, как в клетках M. leprae, произошло и в других внутриклеточных паразитах и патогенах. Пример организмов со значительно уменьшившимся числом функциональных генов показывает, что при изменении образа жизни значительная доля всех генов может стать ненужной.
Исчезновение отдельных генов, групп функционально связанных генов или даже более широкого набора генов у определенного вида имеет важные последствия для будущей эволюции его потомков. Поскольку в разрушающихся генах накапливается множество дефектов, их инактивация обычно является необратимым процессом. Потеря функции гена – это улица с односторонним движением. Функция потеряна, и ее уже не вернуть. Как новые виды ледяных рыб не смогут больше иметь и использовать гемоглобин, так и потомки S. kudriavzevii не смогут питаться галактозой. Потеря генов и нарушение их функций накладывают ограничения на развитие эволюционной ветви.
Используй, или потеряешь – нерушимое правило, обусловленное тем, что естественный отбор действует только в настоящем времени, он не умеет строить планы на будущее. Оборотная сторона этого правила заключается в том, что при смене условий, пусть даже в отдаленном будущем, те виды, которые утратили какие‑то гены, не смогут воспользоваться ими для адаптации к новым условиям. Возможно, это один из важных факторов, объясняющих расцвет или исчезновение видов. Не стоит забывать, что, по мнению биологов, более 99 % некогда существовавших видов организмов на настоящий момент вымерли.
Причина или следствие?
Широкое распространение ископаемых генов открывает большие возможности для изучения эволюции. Но, кроме того, оно поднимает вопрос о причинно‑следственной связи. Является ли отмирание генов причиной эволюционных изменений, вызванных естественным отбором, или это следствие – побочный эффект естественного отбора каких‑то других признаков? По‑видимому, возможно и то и другое – в зависимости от конкретных условий. Я поясню это на примере недавно образовавшихся ископаемых генов у цветковых растений и у человека; в первом случае образование ископаемых генов является, скорее всего, причиной, а во втором – следствием.
У растений окраска цветов часто играет важную роль в привлечении опылителей, особенно пчел и птиц. Известно множество хорошо изученных примеров эволюционных изменений у животных‑опылителей. Легко себе представить, что изменение климата или количества опылителей может повлиять на отбор окраски цветов. Более того, поскольку цветы, снабжающие нектаром пчел и колибри, могут посещать и другие – непрошеные – гости, отбору подвергаются и другие признаки цветка, что позволяет настроить его на различных опылителей. Например, растения, опыляемые птицами, обычно производят больше нектара и имеют цветы вытянутой формы, тогда как растения, опыляемые пчелами, производят меньше нектара и имеют более широкие и плоские цветки.
Раньше у ипомеи, относящейся к семейству вьюнковых, цветы были синими или лиловыми. Эту группу растений обычно опыляют пчелы, однако один вид, Ipomoea quamoclit, имеет красные цветы и опыляется колибри. По‑видимому, красный цвет цветка является адаптацией, способствующей привлечению птиц.
Красный, синий или лиловый цвет цветков ипомеи определяется составом ферментов. Из одного и того же исходного вещества разные наборы ферментов производят либо синий и лиловый, либо красный пигмент. Недавно Ребекка Зуфолл и Марк Раушер из Университета Дьюка (Северная Каролина) показали, что у I. quamoclit, обладающей красными цветками, произошло разрушение метаболического пути синтеза синего и лилового пигментов. Один из генов этого пути полностью испорчен, а второй изменился до такой степени, что может участвовать только в синтезе красного пигмента.
Поскольку появление красной окраски цветков ипомеи является адаптацией, причем, по‑видимому, напрямую связанной с модификацией двух конкретных ферментов, инактивация генов в этом примере, скорее всего, стала причиной эволюционных изменений. Кажется более вероятным, что естественный отбор напрямую способствовал инактивации ферментов для синтеза синего и лилового пигментов и эволюции красной окраски, чем то, что инактивация ферментов стала побочным результатом отбора какого‑то другого признака.
Однако чаще инактивация и разрушение генов являются следствием ослабления давления со стороны естественного отбора, и разрушение генов – это лишь один из заключительных этапов в целой серии изменений. Возможно, именно это случилось с генами опсинов, о которых я уже рассказывал раньше, и это же произошло в удивительном случае, связанном с возникновением ископаемого гена в человеческой линии после того, как предки людей и шимпанзе отделились от общего предка.
Человеческий ископаемый ген, о котором я хочу рассказать, называется MYH16. В последовательности гена MYH16 у человека отсутствуют два основания, что нарушает правильное считывание текста (соответствующий участок последовательности представлен на рис. 5.4; место отсутствующих оснований обозначено звездочками). У шимпанзе, горилл, орангутанов и макак этот ген остался без изменений.

Рис. 5.4. Образование ископаемого гена у человека. Показан короткий фрагмент гена миозина MYH16. У человека произошла делеция двух оснований (звездочки), нарушившая считывание кода и вызвавшая ослабление двух челюстных мышц, сильно развитых у наших родственников – человекообразных обезьян. Рисунок Джейми Кэрролл на основании данных из статьи H. H. Stedman et al., 2004, Nature 428:416.
У других приматов белок MYH16 образуется в некоторых мышцах, особенно в височной мышце, охватывающей большую поверхность височной части черепа с обеих сторон. Эта мышца участвует в движениях нижней челюсти при пережевывании пищи. У человека эта часть лица и эта мышца гораздо менее развиты, чем у горилл, шимпанзе и макак. Продукт гена MYH16 представляет собой белок миозин, являющийся составным элементом мышечных волокон. Волокна височной мышцы человека значительно тоньше, чем у других приматов, а более тонкие волокна образуют более слабую мышцу.
Интересная связь между мутацией белка, влияющего на размер мышечных волокон и мышц, и эволюционными изменениями височной мышцы вызывает вопрос, была ли мутация причиной ослабления мышцы или побочным результатом ослабления мышцы, вызванного другими причинами. Однозначно ответить на этот вопрос трудно, но давайте познакомимся с некоторой дополнительной информацией по этому поводу. Известно, что мутации в этом классе белков могут оказывать значительное влияние на состояние мышц. Если примат с тяжелой челюстью (как у человекообразных обезьян и наших с вами предков) сразу лишится мышцы, он не сможет жевать. Если мутация MYH16 сыграла роль в эволюции височной мышцы, скорее всего, мышечная масса сокращалась поэтапно. Мне кажется, что наиболее вероятный сценарий потери функции гена MYH16 напоминает историю с глобинами у ледяных рыб. Это означает, что мышца ослабла в результате других генетических причин, а превращение гена MYH16 в ископаемый ген, скорее всего, было более поздним событием и объяснялось отсутствием потребности в этом гене.
Дата добавления: 2018-09-22; просмотров: 360; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!