Появление эукариот: союз двух непохожих родителей?
Я верю, что придет время, хотя я и не доживу до тех пор, когда у нас будет истинное генеалогическое древо для каждого царства живых организмов.
Чарльз Дарвин Из письма Т. Хаксли, 26 сентября 1857 г.
Со времен Дарвина наше понимание устройства природы значительно продвинулось вперед. Раньше всю живую природу подразделяли лишь на царство растений и царство животных. Эта двоичная система существовала со времен Аристотеля и была формально описана Карлом Линнеем в 1735 г. В 1866 году Эрнст Геккель, осуществивший замечательное исследование простейших организмов, добавил к этой классификации третье царство – царство протистов. Бактерии и грибы были включены в классификацию в качестве полноправных царств только в начале XX в.
Кроме распределения по пяти царствам признавалось и разделение более высокого порядка, основанное на фундаментальном различии клеток, обнаруженных у представителей разных царств. В 1938 году французский биолог Эдуард Шатон предложил называть представителей этих «надцарств» прокариотами и эукариотами в зависимости от присутствия или отсутствия ядра в их клетках. В эти надцарства попадали все известные формы живых организмов, но лишь до тех пор, пока Карл Воуз не начал изучать гены организмов, обнаруженных Томом Броком в Йеллоустоне.
Воуз считал, что таксономия бактерий слишком запутанна и что для определения эволюционных связей между видами требуются более объективные показатели, чем их морфология или физиология. Он обратился к изучению молекул. Как только анализ белковых последовательностей стал выявлять сходство и различия между белками, общими у разных видов, многие ученые (такие как Фрэнсис Крик, Эмиль Цукеркандль и Лайнус Полинг) признали, что можно строить генеалогическое древо живых организмов на основании последовательностей ДНК, РНК и белков. В основе этого подхода лежала очень простая идея. Участки в последовательности ДНК, РНК или белков, различающиеся у одних групп видов, но одинаковые у других, указывают на степень родства этих видов. Так же как семейное древо строится на основе степеней родства, генеалогическое древо живых организмов строится на основе генетического сходства. Однако, как я объясню далее, иногда случаются такие союзы, которые в немалой степени запутывают семейную генеалогию.
Для создания генеалогического древа бактерий Воуз использовал имеющийся в изобилии тип молекул РНК. Однако, когда он включил в свою схему термофильные метанобразующие организмы, оказалось, что «эти „бактерии“ похожи на типичные бактерии не больше, чем на эукариотическую цитоплазму». Он предположил, что существует третье надцарство, куда следовало бы отнести эти организмы, приспособленные к жизни в различных экстремальных условиях. Поскольку считается, что именно такие условия существовали на Земле в самом начале ее истории, это царство могло бы считаться первичным, или процарством, так что Воуз предложил назвать это надцарство «архебактериями». Позднее архебактерии стали называть археями, отчасти чтобы не путать их с бактериями, а «надцарства» переименовали в «домены».
Хотя разделение всех форм жизни на три домена – эукариоты, архей и бактерии – закрепилось в науке, в родственных связях между ними оказалось не так‑то просто разобраться. По представлениям Дарвина каждая ветвь генеалогического древа жизни соответствовала образованию нового вида. Однако в мире микробов, с которым Дарвин не был знаком, происходят события, нарушающие стандартный способ ветвления. Микробы могут обмениваться генами, а некоторые микробы живут внутри других организмов‑хозяев (так называемый эндосимбиоз). И то и другое способствует переносу генов между очень отдаленными видами организмов и нарушает «правильное» ветвление генеалогического древа. Чтобы понять связь между эукариотами, археями и бактериями, биологи должны установить происхождение множества генов, далеко не все из которых могут иметь признаки семейного сходства.
Например, в ранних исследованиях было обнаружено удивительное сходство между некоторыми молекулами архей и молекулами эукариотов. Белки, которые археи используют для упаковки ДНК в хромосомы, транскрипции ДНК и расшифровки генетической информации, настолько похожи на соответствующие белки эукариот, что высказывалось предположение о том, что эукариоты произошли от каких‑то видов архей. Это сходство, в частности, присутствует в коротких специфических последовательностях («подписях») в составе белков, которые являются общими для некоторых архей и эукариот, но отсутствуют в других группах организмов. Например, в одном из бессмертных белков, участвующих в расшифровке генетической информации, есть короткая вставка из и аминокислотных остатков. В таблице 3.2 представлена последовательность этой вставки у различных эукариот и архей.
Таблица 3.2. Последовательность вставки

Присутствие этой последовательности в двух доменах и ее отсутствие в третьем логично было бы объяснить тем, что архей и эукариоты более близкие родственники друг другу, чем бактериям. Если строить древо жизни исходя из этого соображения, то получается, что общий предшественник всех трех доменов (так называемый последний общий универсальный предшественник, LUCA – от англ. last universal common ancestor) в какой‑то момент дал начало двум доменам – бактериям и археям, а позднее от ветви архей отделились эукариоты. В этом случае древо жизни имело бы такой вид, как изображено на рис. 3.4.

Рис. 3.4. «Традиционная» форма древа жизни. Все домены возникают как ветви на стволе дерева. Рисунок Джейми Кэрролл.
Однако в результате секвенирования целых геномов архей и бактерий довольно неожиданно обнаружилось, что очень многие гены архей похожи на соответствующие гены бактерий. Затем по мере расшифровки все большего числа эукариотических геномов стало выясняться, что многие гены эукариот ближе к генам бактерий, чем к генам архей. Возникла ситуация как в известной загадке: «Если твоя сестра приходится тебе тетей, то кто твой отец?» Одним словом, ответить на вопрос о том, кто кому более близкий родственник, оказалось совсем не просто.
Ответ был найден в результате дополнительных исследований. Было замечено, что основное сходство между генами архей и эукариот касается так называемых информационных генов, продукты которых участвуют в удвоении и экспрессии ДНК. А наибольшее сходство между генами эукариот и бактерий обнаружено в операционных генах, отвечающих за метаболизм различных питательных веществ и основных клеточных компонентов. Это могло означать, что эукариоты взяли «ум» (информационные гены) от одного родителя, а «внешность» (операционные гены) – от другого;
Отсюда родилось подозрение, что эукариоты являются плодом смешанного союза – генетического слияния архей и бактерий. Идея о слиянии сильно различающихся видов не является новой. В 1970 году Линн Маргулис[11] предположила, что митохондрии и хлоропласты – две основные производящие энергию клеточные органеллы в эукариотических клетках – происходят из бактерий, живущих внутри эукариот (этот процесс называется эндосимбиозом). Сегодня эта точка зрения широко распространена.
Но правда ли, что эукариоты произошли от союза архей и бактерий? Мария Ривера и Джеймс Лейк из Калифорнийского университета в Лос‑Анджелесе пришли к выводу, что эукариоты и в самом деле имеют двоякую природу, то есть происходят от родителей, находящихся на разных ветвях древа жизни. Ривера и Лейк анализировали геномы бактерий, архей и эукариотов в поисках групп генов, имеющихся абсолютно во всех основных подразделах этих трех доменов, а также генов, отсутствующих в одном, двух, трех и т. д. из этих подразделов. Всесторонний анализ распределения общих генов показывает, что геном эукариот является продуктом слияния геномов каких‑то видов архей и каких‑то видов бактерий. Симбиотические взаимоотношения достаточно часто встречаются у живущих вместе организмов (так, Thermus aquaticus из горячих источников в Йеллоустоне получает энергию от фотосинтезирующих цианобактерий, которые и придают окраску почве вокруг источников), и время от времени это приводит к эндосимбиозу, поэтому возможным объяснением происхождения эукариот является слияние геномов эндосимбионта и его хозяина. Таким образом, основание древа жизни – это не точка, а круг, из которого наше древо восходит и разветвляется (рис. 3.5).
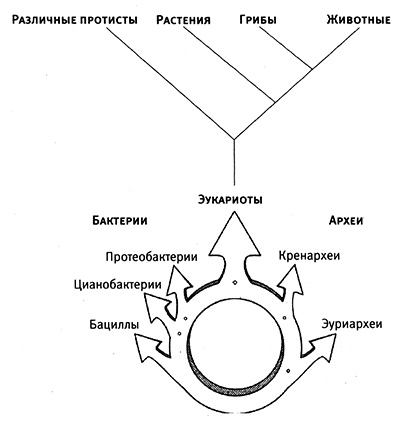
Рис. 3.5. Новое изображение филогенетического древа эукариот. Анализ ДНК показывает, что эукариоты появились в результате слияния каких‑то видов архей с какими‑то видами бактерий. Таким образом, основанием древа является круг, а не точка. Из статьи M. Rivera, J. Lake, 2004, Nature 431:152 (с изм.).
Поэтому, если вам повезет и вы посетите замечательный парк в Йеллоустоне, не отворачивайтесь от кипящего в горячих источниках неприятно пахнущего супа и сдержите свое отвращение при виде ярко окрашенной тины. Это невежливо по отношению к родственникам, пусть даже самым дальним. Обдумайте тот забавный факт, что вас с представителями этого сообщества роднят сотни общих генов. И именно в этом сообществе, когда‑то безумно давно, возможно, где‑то в глубоководных термальных источниках, из столба метана возник предок всех существующих на Земле живых организмов.
Конечно, если бы естественный отбор сводился к поддержанию порядка в очень жестких пределах, жизнь была бы однотипной и неизменной и не имела бы того бурного разнообразия форм, которое мы видим вокруг и в летописи окаменелостей за последние 3 млрд лет. Данные, приведенные в табл. 3.1, говорят о том, что геномы разных видов очень сильно различаются. Если оставить в стороне 500 общих бессмертных генов, геномы различаются как по количеству генов, так и по последовательностям. Различие в количестве генов говорит о том, что в ходе эволюции должны были возникать новые гены. Они и вправду возникали – это созидательное измерение эволюции станет темой следующей главы. Но раз гены рождаются, они могут и умирать. Это тоже верно – гены умирают, и о том, что это значит для эволюции, мы поговорим в пятой главе.

Обезьяна колобус в парке Кибале (Уганда). Эта обезьяна способна находить более питательные листья и переваривать их благодаря двум эволюционным приобретениям – полноценному цветовому зрению и специфической системе пищеварения, напоминающей систему пищеварения жвачных животных. Фотография Кагана Секерсиоглу.
Глава 4
Сделать новое из старого
Сохраняй старое, но познавай новое.
Китайская пословица
В парке Кибале время завтрака.
Высоко под куполом леса стая нарядных черно‑белых колобусов перебирается на новое место в поисках пищи. Можно предположить, что рацион питания этих обитателей густых дождевых лесов Уганды будет бесконечно разнообразен. Однако колобусы не обращают никакого внимания на изобилие зелени и выбирают растения с красноватыми листьями. Единственный вид обезьян, у которых нет большого пальца (их название происходит от греческого слова kolobus, что означает «искалеченный» или «обрубленный»), колобусы направляют в рот гибкие ветви с молодыми листьями, ловко работая своими четырьмя пальцами.
Чуть ниже, ближе к земле, несколько особей из пяти сотен обитающих здесь шимпанзе с криками пробираются к фиговым деревьям. Они тоже иногда жуют листья, но по‑настоящему их интересуют только спелые красные или желтые плоды. Шимпанзе не любят неспелые фрукты, поэтому они срывают несколько плодов на пробу, удобно усевшись на ветке.
Для поиска пищи колобусы и шимпанзе пользуются своим полноценным цветовым зрением, которое отсутствует у млекопитающих, не относящихся к приматам. Все обезьяны Старого Света (африканские и азиатские), включая человекообразных обезьян, обладают трихроматическим зрением, которое позволяет им различать цвета во всем видимом спектре – от синего до красного. Большинство других млекопитающих имеют дихроматическое зрение, то есть могут видеть синий и желтый цвет, но не отличают красный от зеленого. Обладание трихроматическим зрением играет очень важную роль, поскольку наиболее нежными и питательными являются молодые листья, а в тропиках более чем у половины растений они окрашены в красный цвет. Используя это различие в цвете, невидимое для других листоядных животных, приматы собирают самые питательные листья.
Переваривать лиственный завтрак колобусам помогает уникальная способность, которой обладают только они и их ближайшие родственники – в отличие от шимпанзе и всех остальных приматов. Колобусы – жвачные животные! Они могут питаться листьями и травой, поскольку располагают необычайно большим желудком с множеством камер. Взрослые особи с массой тела около 15 кг съедают ежедневно около 3 кг листьев, и от этого у них образуется весьма заметный животик. Бактерии в кишечнике колобуса помогают переварить растительную пищу, пока она медленно продвигается по пищеварительному тракту, а особые ферменты позволяют расщепить важнейшие питательные вещества, высвобождаемые бактериями.
Зрение и пищеварительная система этих приматов заставляют вновь задаться одним из величайших вопросов биологии: как у живых организмов появляются новые признаки? В этой главе мы рассмотрим механизмы, которые позволяют живым организмам приобретать новые способности и развивать старые таланты. Мы узнаем о том, как новые функции и новые гены возникают из «старых» генов. Я расскажу, как случайное удвоение гена обеспечивает запчасти для эволюции новых функций и как эти новые и старые части генома настраиваются для адаптации к определенному образу жизни.
Я мог бы привести множество примеров нововведений и модификаций. Однако я в основном буду рассказывать о происхождении и эволюции цветового зрения, и для этого есть несколько веских причин. Во‑первых, преимущества этой способности очевидны. Во‑вторых, зрительные системы животных, обитающих в самых разных условиях (в океанах, саваннах, лесах, пещерах, под землей и т. д.), поразительно адаптированы именно к этим конкретным условиям. В‑третьих, мы достаточно хорошо знаем биологию и физику процессов, лежащих в основе цветового зрения, и поэтому можем оценить большие и малые различия в зрительных способностях живых организмов и в том, какие цвета они могут воспринимать. Мы знаем, что существует широкий ультрафиолетовый диапазон спектра, который невидим для человека, но птицы, насекомые и многие другие живые существа используют его для поиска пищи, партнеров и сородичей. В‑четвертых, гены, отвечающие за цветовое зрение, возможно, изучены лучше, чем гены, отвечающие за какие‑либо другие функции. Все вместе дает нам колоссальный объем знаний, позволяющий связать различия между определенными генами с различиями в экологии и эволюции видов.
Новые научные данные, с которыми мы познакомимся в этой главе, служат ясным и прямым доказательством трех аспектов эволюции – естественного отбора, полового отбора и происхождения видов путем наследования с изменениями. Эти три аспекта отчетливо проявляются при анализе хода эволюции по генетическим данным. Чтобы проследить этапы эволюции, нам придется идентифицировать важные различия между отдельными видами, определить, когда эти различия возникли, и описать, как те или иные изменения на уровне ДНК влияют на природные особенности организмов. Для этого нам нужны два типа данных, полученных из ДНК. Во‑первых, чтобы узнать, когда и в каких видах произошли изменения, мы обратимся к недавно обнаруженным уникальным последовательностям ДНК, что позволит нам получить четкую картину взаимосвязей между видами. Во‑вторых, нам придется тщательно проанализировать ДНК‑код тех генов, которые отвечают за цветовое зрение. В последовательностях этих генов запечатлены следы естественного отбора.
Журналист Рекс Дальтон писал о мире, который видят птицы, но не видим мы: «Если вы хотите узнать мысли животных, посмотрите на мир их глазами». Я начну с того, что объясню, как животные различают цвета, а затем мы с вами проследим ход эволюции при помощи этих самых глаз.
Рассматриваем радугу
Зрение человека уникально. Мы различаем цвета и их оттенки благодаря группе тонко настраиваемых молекул, которые распознают свет в клетках нашей сетчатки и посылают сигналы в головной мозг. У других животных набор этих молекул иной, или они настроены на восприятие других частей спектра (а возможно, и то и другое). Чтобы понять, что же видим мы и другие животные, нам нужно кое‑что узнать о свете и цвете, о молекулах, которые детектируют свет, и о клетках глаза, собирающих цветовое изображение.
Наши глаза очень чувствительны к тому диапазону спектра, который мы, с нашей антропоцентрической точки зрения, называем видимым светом. На самом деле это узкая полоска в широком спектре электромагнитного излучения. Белый цвет является результатом смешения всех цветов видимого диапазона спектра, простирающегося от фиолетового до синего, зеленого, желтого, оранжевого и красного. Каждому цвету радуги соответствует определенная длина волны: фиолетовый свет имеет длину волны около 400 нанометров (нм), а красный – около 700 нм (рис. 4.1).

Рис. 4.1. Спектр видимого света. Фиолетовый, синий, зеленый и красный свет – это свет с разной длиной волны (измеряется в нанометрах, нм). Цветовое зрение человека настроено на восприятие света с четырьмя разными длинами волн (указаны стрелками,) с помощью четырех разных белков опсинов. Рисунок Лианн Олдс.
Цвет предметов зависит от длины волн поглощенного или отраженного света, а это, в свою очередь, определяется молекулярной структурой этих предметов. Например, траву мы видим зеленой, поскольку она поглощает свет во всем диапазоне видимого спектра, кроме зеленого; зеленый свет с длиной волны 520 нм трава отражает. Небо кажется нам голубым, поскольку атмосфера рассеивает волны солнечного света, большинство которых голубые. Оставшаяся часть солнечного света кажется желтой, поскольку так выглядит белый цвет, если из него вычесть синий. Закат мы видим оранжевым, поскольку при приближении солнца к линии горизонта свет проделывает более долгий путь через атмосферу, прежде чем достичь наших глаз, и по дороге рассеивается большая часть синего, в результате чего остается оранжевый цвет. В солнечном излучении есть и более коротковолновая составляющая, которую мы не видим, в частности ультрафиолетовое излучение, рассеиваемое атмосферным озоном. Та часть ультрафиолетового излучения, которая все же достигает земли, поглощается кожей и вызывает ожоги у таких бледнокожих представителей человечества, как я, а тех, кому повезло больше, чем мне, покрывает красивым загаром. Свет с большей длиной волны, как тот, что выделяется в виде тепла от горящего огня, называют инфракрасным излучением. Его тоже нельзя увидеть невооруженным глазом, но можно зарегистрировать с помощью очков ночного видения – специального устройства с детекторами инфракрасного излучения, которые реагируют на тепло.
Восприятие цвета начинается тогда, когда свет с определенной длиной волны сталкивается со зрительными пигментами в сетчатке глаза. Эти пигменты состоят из белка опсина и небольшой молекулы хромофора, которая у человека является производным витамина A. Чувствительность зрительного пигмента к свету определяется последовательностью белка опсина и его взаимодействием с хромофором. Это взаимодействие обеспечивает тонкую спектральную настройку, в результате которой каждый зрительный пигмент настраивается на свет с определенной длиной волны. У человека есть три вида зрительных пигментов, чувствительных к коротким, средним и длинным световым волнам. Их называют коротковолновыми, средневолновыми и длинноволновыми опсинами (или опсинами SWS, MWS и LWS). Опсин SWS настроен на длину волны 417 нм (синий цвет), опсин MWS – на 530 нм (зеленый цвет), а опсин LWS – на 560 нм (красный цвет), и вместе они обеспечивают нам цветовое зрение. Четвертый пигмент, родопсин (497 нм), используется в основном при слабом освещении. Свет с длиной волны меньше 400 нм (ультрафиолетовая область) или больше 700 нм (инфракрасная область) невидим для человека, но, как я расскажу позднее, многие животные могут видеть в ультрафиолетовом диапазоне спектра.
Глаз человека снабжен двумя типами фоторецепторных клеток – палочками и колбочками, которые получили свое название благодаря форме своих внешних частей, содержащих зрительные пигменты (рис. 4.2).

Рис. 4.2. Восприятие цвета сетчаткой человеческого глаза. Свет попадает в глаз через хрусталик, а затем достигает сетчатки. В сетчатке есть два типа фоторецепторных клеток: палочки, которые позволяют видеть при слабом освещении, но не создают цветовых изображений, и колбочки, содержащие опсины одного из трех видов (у человека), настроенные на восприятие красного (К), зеленого (3) или синего (С) цвета. Возбуждение палочек и колбочек интегрируется нейронами, которые соединяются в зрительный нерв. Далее зрительный нерв проводит сигнал к зрительным центрам головного мозга. Рисунок Лианн Олдс.
В целом палочки обладают очень высокой чувствительностью к свету и наиболее интенсивно используются ночью и при слабом освещении, но они не способны различать длину световой волны, поэтому ночью мы плохо различаем цвета. Колбочки используются при ярком свете, и именно с их помощью мы воспринимаем цвет.
Когда свет попадает на хромофор, он вызывает серию очень быстрых изменений зрительного пигмента. За тысячные доли секунды пигмент переходит в возбужденное состояние, что активизирует фоторецепторные клетки. В конечном итоге сигнал фоторецепторов сетчатки достигает зрительных центров коры головного мозга. Чтобы изображение было воспринято как цветное, должны возбудиться как минимум два типа колбочек. Цвет «видимых» мозгом предметов определяется относительным уровнем возбуждения каждого типа колбочек. Если в сетчатке присутствует лишь один тип колбочек, все будет видеться в серой цветовой гамме.
Каждый опсин кодируется отдельным геном. У шимпанзе и других обезьян имеются те же три опсина, что и у человека (SWS, MWS и LWS). Однако у большинства других млекопитающих есть лишь два опсина (и два соответствующих гена), а вот у птиц и рыб – четыре опсина и более. Таким образом, очевидно, что число генов опсинов изменялось в ходе эволюции животных. Эволюция генов опсинов шла через удвоение генов, которое является одним из важных механизмов повышения информационного содержания ДНК. В этом процессе последовательность существующего гена удваивается, а затем пути «старой» и «новой» копии расходятся, и они превращаются в отдельные гены с разными функциями.
Мы знаем, что с опсинами это происходило в ходе эволюции позвоночных животных, но нам хотелось бы знать больше, а именно, какие конкретно животные приобрели (или потеряли) гены опсинов. Недостаточно просто сказать, что два вида различаются между собой: чтобы проследить этапы эволюции, нужно иметь перед глазами картину того, что было до и что было после интересующего нас события. Чтобы узнать истинную историю генов опсинов, нам нужно хорошо представлять себе родственную связь между разными группами животных. Знание родственных связей между организмами позволяет понять направление эволюции конкретного признака и определить состояние этого признака и соответствующего гена у общих предков. Например, если два родственных вида имеют общий признак или общий ген, скорее всего, это не является совпадением, а объясняется тем, что их общий предок также обладал этим признаком или геном. Определить происхождение признака невозможно без знания генеалогии видов. Чтобы представить себе, как была установлена генеалогия видов, имеющих отношение к появлению нашего цветового зрения, нужно хотя бы в общих чертах понять, как сегодня, в эпоху ДНК, устанавливают родственные связи между видами. Я расскажу о недавно обнаруженных и очень важных последовательностях ДНК, которые позволяют реконструировать историю видов с небывалой доселе ясностью и достоверностью.
От молекул к деревьям
С тех пор как Дарвин предположил, что эволюционное развитие организмов можно представить в виде ветвящегося дерева, биологи все время рисуют подобные деревья для изображения родственных связей между ныне живущими и когда‑то жившими организмами. Раньше подобные деревья строились на основании внешних признаков современных организмов или ископаемых остатков древних организмов. Однако довольно часто выяснялось, что внешнее сходство или его отсутствие уводит не в ту сторону или по меньшей мере вызывает споры между биологами, поэтому практически для всех основных групп организмов было создано несколько генеалогических деревьев.
Несколько десятилетий назад для определения степени родства между видами ученые начали использовать последовательности биологических молекул. Гены наследуются, и это означает, что последовательности генов и кодируемых ими белков передаются от родителей детям. Происходящие в этих последовательностях изменения также передаются потомкам на всем долгом пути эволюции. Таким образом, степень сходства ДНК может служить показателем близости родства между видами.
Теоретически для построения генеалогического древа биологи имеют в своем арсенале тысячи последовательностей ДНК или белков. На практике для выбора генов для построения древа используют несколько критериев отбора (в частности, стоимость анализа), так что обычно из многих генов‑кандидатов выбирают лишь один или несколько. На основании выбранных последовательностей генов с помощью сложного математического и статистического аппарата строят генеалогическое древо. Это древо должно как можно точнее соответствовать имеющимся экспериментальным данным. Так как последовательности генов состоят всего из четырех знаков (A, C, G и T), при недостаточном количестве данных родство между организмами может быть определено неточно. Не следует думать, что с помощью любого гена можно получить один и тот же правильный или однозначный ответ. Поэтому один из способов проверить, насколько верно выстроено древо, – попытаться воспроизвести его на основании другой последовательности или более широкого круга последовательностей.
К счастью, был разработан совершенно новый метод определения родственных связей между видами. Он также основан на анализе ДНК, но учитывает не степень сходства последовательностей в целом, а наличие или отсутствие определенных маркеров в специфических участках ДНК. Эти маркеры возникают в результате случайного встраивания мусорной ДНК рядом с генами. Достаточно часто в ДНК встречаются специфические последовательности, называемые длинными диспергированными повторами (LINE) и короткими диспергированными повторами (SINE). Если такие повторы встраиваются в последовательность, они в ней остаются, поскольку не существует механизма их активного удаления. Последовательность этих элементов в составе ДНК является характерной для данного вида и передается по наследству всем потомкам. Это замечательный инструмент для построения генеалогического древа. Случаи встраивания подобных последовательностей встречаются очень редко, поэтому их присутствие в одном и том же участке ДНК двух разных видов может объясняться исключительно происхождением этих видов от общего предка. На том же принципе наследования маркеров в ДНК основано определение отцовства у людей. Анализ распределения нескольких маркерных элементов ДНК, возникших в разное время у разных организмов, дает биологам достаточно данных, чтобы абсолютно точно устанавливать родственные отношения между видами.
Мы вкратце ознакомились с сутью процесса построения генеалогического древа, но давайте не будем ставить точку. Мне хочется, чтобы, читая эту книгу, вы в полной мере оценили природу и качество имеющихся доказательств эволюции. Чтобы поверить и понять, нужно увидеть. Поэтому давайте посмотрим, как с помощью повторов SINE было построено такое важное генеалогическое древо, как древо человека и других приматов, а затем при помощи этого древа раскроем тайну происхождения цветового зрения.
Прежде всего при построении древа нужно обнаружить набор повторов SINE. Последовательность генома человека известна, что облегчает задачу: мы можем выбирать из тысяч коротких диспергированных повторов. В соответствующих участках ДНК других приматов ищут те же самые повторы.
Большинство SINE состоят примерно из 300 пар оснований, поэтому, если в ДНК какого‑то организма содержится такой же SINE, как в ДНК человека, этот фрагмент ДНК будет на 300 пар оснований длиннее, чем в случае отсутствия SINE. На рис. 4.3 представлены данные, полученные Абдель‑Халимом Салемом и его коллегами из университетов Луизианы и Юты.

Рис. 4.3. Типирование ДНК и эволюция гоминидов. Наличие определенных повторов SINE в ДНК разных видов организмов определяют по яркой полосе на соответствующей дорожке в геле. Наличие одинаковых повторов у представителей разных видов указывает на их эволюционное родство. Из статьи Salem et al., 2003, Proceedings of the National Academy of Science 100:12 787. © 2003 by the National Academy of Sciences, USA.
Наличие или отсутствие SINE определяется по положению полос ДНК в геле, используемом для разделения фрагментов ДНК разного размера. Один из повторов, представленных на рисунке, специфичен только для человека, второй является общим для человека, карликовых шимпанзе (бонобо) и шимпанзе, а третий – для человека, бонобо, шимпанзе, горилл, орангутанов и сиамангов (сростнопалых гиббонов). Среди 100 с лишним повторов были выявлены такие, которые являются общими для разных групп приматов, в том числе для всех шести исследованных видов человекообразных обезьян, а также лишь для пяти, четырех и т. д. видов человекообразных. Также были выявлены повторы, которые являются общими лишь для бонобо и шимпанзе, и уникальные для человека, бонобо или шимпанзе. Ни один из исследованных повторов не был обнаружен в геноме ночных (совиных) обезьян. Количество повторов, являющихся общими для разных видов, отражает степень родства этих видов, и может служить для построения генеалогического древа (рис. 4.4).

Рис. 4.4. Эволюционное древо гоминоидов. Эволюционное родство между гоминидами и обезьянами, установленное на основании анализа ДНК. На основании данных из Salem et al., 2003, Proceedings of the National Academy of Science 100:12 787.
Из этого древа следует, что ближайшими родственниками человека являются шимпанзе, ближайшими родственниками шимпанзе – бонобо, а ветви горилл и орангутанов отделились от древа до появления последнего общего предка человека и шимпанзе. Достоверность этой информации не подлежит сомнению.
Теперь давайте с помощью этого древа займемся изучением происхождения цветового зрения и опсинов.
Дата добавления: 2018-09-22; просмотров: 302; Мы поможем в написании вашей работы! |

Мы поможем в написании ваших работ!